–Э–∞ –Р-—Б–∞–є—В–µ, –≤ "–Ш–Ј–±–µ-–Ф–µ–±–∞—В–љ–µ", —Б –≤–µ—Б–љ—Л 2001 –≥–Њ–і–∞ —А–∞–Ј–≥–Њ—А–µ–ї—Б—П –і–Є—Б–њ—Г—В —Б –≤–µ—А—Г—О—Й–Є–Љ–Є –њ–Њ —В–µ–Љ–µ —Н–≤–Њ–ї—О—Ж–Є–Є –Є –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П (вАЬ–Ш–Ј–±–∞-–Ф–µ–±–∞—В–љ—ПвАЭ, —В–µ–Љ–∞ вДЦ 39). –Т —Б–њ–Њ—А–∞—Е –∞—В–µ–Є—Б—В–∞–Љ –±—Л–ї –±—А–Њ—И–µ–љ (–Ш–Ь–•–Ю –≥–і–µ-—В–Њ —Б–њ—А–∞–≤–µ–і–ї–Є–≤–Њ) —Г–њ—А—С–Ї, —З—В–Њ –≤—Б–µ –њ—А–Є–≤–µ–і–µ–љ–љ—Л–µ –і–Њ —Н—В–Њ–≥–Њ —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ—Л–µ –њ—А–Є–Љ–µ—А—Л –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П , –Ї–∞–Ї –Є–Ј –ґ–Є–≤–Њ–є –њ—А–Є—А–Њ–і—Л, —В–∞–Ї –Є –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л–µ, –љ–µ –њ–Њ–Ї–∞–Ј—Л–≤–∞—О—В —В–Њ–љ–Ї–Њ—Б—В–µ–є –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П вАУ –љ–∞–Ї–Њ–њ–ї–µ–љ–Є–µ –Є–Ј–Љ–µ–љ–µ–љ–Є–є –≤ –Ї–∞—А–Є–Њ—В–Є–њ–µ - –Є –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–љ—Г—О —Н—В–Є–Љ —З–∞—Б—В–Є—З–љ—Г—О –Є–ї–Є –њ–Њ–ї–љ—Г—О –≤–Є–і–Њ–≤—Г—О –Є–Ј–Њ–ї—П—Ж–Є—О.–Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –≤–Њ–Ј—А–∞–ґ–µ–љ–Є—П –±—Л–ї–Є –Ї —Б–∞–Љ–Є–Љ –њ—А–Є–Љ–µ—А–∞–Љ –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П, –≥–і–µ –њ—А–Є–≤–Њ–і–Є–ї–Є—Б—М –≤–Є—А—Г—Б—Л, —А–∞—Б—В–µ–љ–Є—П, —З–µ—А–≤—П–Ї–Є, –Љ—Г—Е–Є, —Б–∞–ї–∞–Љ–∞–љ–і—А—Л –Є –і—А—Г–≥–Є–µ "–њ—А–Њ—Б—В—Л–µ" (–≤ –њ–Њ–љ–Є–Љ–∞–љ–Є–Є –≤–µ—А—Г—О—Й–Є—Е) –Њ—А–≥–∞–љ–Є–Ј–Љ—Л, –Ї–Њ—В–Њ—А—Л–µ "–љ–µ–і–Њ—Б—В–Є–ґ–Є–Љ–Њ –і–∞–ї–µ–Ї–Є" –Њ—В —З–µ–ї–Њ–≤–µ—З–µ—Б–Ї–Њ–≥–Њ —Б–Њ–≤–µ—А—И–µ–љ—Б—В–≤–∞ –Є —Б–ї–Њ–ґ–љ–Њ—Б—В–Є –≤–љ—Г—В—А–µ–љ–љ–µ–≥–Њ —Г—Б—В—А–Њ–є—Б—В–≤–∞.–≠—В–Њ –і–∞–≤–∞–ї–Њ –њ–Њ–≤–Њ–і –Њ–њ–њ–Њ–љ–µ–љ—В–∞–Љ —Б–Њ–Љ–љ–µ–≤–∞—В—М—Б—П –≤ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є —Н–≤–Њ–ї—О—Ж–Є–Є –Є –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П —В–∞–Ї–Є—Е –љ–µ–њ—А–Њ—Б—В—Л—Е –Є –≤—Л—Б–Њ–Ї–Њ–Њ—А–≥–∞–љ–Є–Ј–Њ–≤–∞–љ–љ—Л—Е —Б—Г—Й–µ—Б—В–≤, –Ї–∞–Ї –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є–µ (–≤ —В.—З. —З–µ–ї–Њ–≤–µ–Ї–∞).
–Э–∞–і–Њ –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ –≤ –њ—А–Є—А–Њ–і–µ (—В–∞–Ї–Є–µ –і–∞–љ–љ—Л–µ –Љ–љ–µ –њ–Њ–њ–∞–і–∞–ї–Є—Б—М) –љ–∞—Б—З–Є—В—Л–≤–∞–µ—В—Б—П 3-4 –Љ–ї–љ.—А–∞–Ј–ї–Є—З. –≤–Є–і–Њ–≤ –ґ–Є–≤–Њ—В–љ—Л—Е. –Я–Њ–љ—П—В–љ–Њ, —З—В–Њ –і–Њ –Є–Ј—Г—З–µ–љ–Є—П —Н–≤–Њ–ї—О—Ж. –њ—Г—В–µ–є –≤—Б–µ–≥–Њ –≤–Є–і–Њ–≤–Њ–≥–Њ –Љ–љ–Њ–≥–Њ–Њ–±—А–∞–Ј–Є—П , –љ–∞ —Г—А–Њ–≤–љ–µ –њ–Њ—Б–ї–µ–і–љ–Є—Е –і–Њ—Б—В–Є–ґ–µ–љ–Є–є –љ–∞—Г–Ї–Є, —Г –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–µ–є –њ–Њ–Ї–∞ –љ–µ –і–Њ—И–ї–Є —А—Г–Ї–Є. –Ф–ї—П —Н—В–Њ–≥–Њ –љ–µ —Е–≤–∞—В–∞–µ—В —Б—А–µ–і—Б—В–≤ (–Њ—Б–Њ–±–µ–љ–љ–Њ –≤ –†–Њ—Б—Б–Є–Є), –≤—А–µ–Љ–µ–љ–Є, –Ї–≤–∞–ї–Є—Д–Є—Ж. –љ–∞—Г—З–љ. –Ї–∞–і—А–Њ–≤ –Є –њ—А–Њ—З.
–Т—В–Њ—А–Њ–µ –≤–∞–ґ–љ–Њ–µ –Ј–∞–Љ–µ—З–∞–љ–Є–µ - –њ—А–Є—А–Њ–і–∞ –љ–µ –ґ–і—С—В –љ–∞—Б —Б –≥–Њ—В–Њ–≤—Л–Љ–Є, "–Ј–∞–Љ–Њ—А–Њ–ґ–µ–љ–љ—Л–Љ–Є" –і–ї—П –∞—А–≥—Г–Љ–µ–љ—В–Њ–≤ –≤ —Б–њ–Њ—А–µ, –њ—А–Є–Љ–µ—А–∞–Љ–Є –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П "–њ–Њ–і –Ј–∞–Ї–∞–Ј", –љ–∞ –≤—Б–µ —Б–ї—Г—З–∞–Є –ґ–Є–Ј–љ–Є. –Ь–Њ–ґ–љ–Њ —В–Њ–ї—М–Ї–Њ –љ–∞–±–ї—О–і–∞—В—М –Є –Ї–Њ–љ—Б—В–∞—В–Є—А–Њ–≤–∞—В—М —Д–∞–Ї—В –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –љ–∞ –Ї–∞–Ї–Њ–Љ-—В–Њ –µ–≥–Њ –Ї–Њ–љ–Ї—А–µ—В–љ–Њ–Љ —Н—В–∞–њ–µ (—Н—В–∞–њ –Љ–Њ–ґ–µ—В –Є –Њ–њ–µ—А–µ–ґ–∞—В—М –Є –Њ—В—Б—В–∞–≤–∞—В—М –Њ—В —В—А–µ–±—Г–µ–Љ–Њ–≥–Њ –і–ї—П –і–Є—Б–њ—Г—В–∞). –Ф—Г–Љ–∞—О, —Н—В–Њ –њ–Њ–љ—П—В–љ–Њ, –µ—Б–ї–Є —Г—З–µ—Б—В—М, —З—В–Њ –Њ—Б—П–Ј–∞–µ–Љ—Л–µ —Н–≤–Њ–ї—О—Ж. –Є–Ј–Љ–µ–љ–µ–љ–Є—П –Љ–Њ–≥—Г—В —В—А–µ–±–Њ–≤–∞—В—М —В—Л—Б—П—З–Є –ї–µ—В –і–ї—П –Њ–і–љ–Њ–≥–Њ –Љ–∞–ї–µ–љ—М–Ї–Њ–≥–Њ —И–∞–ґ–Ї–∞ –≤–њ–µ—А—С–і.
–Т —В—А–µ—В—М–Є—Е, –≤ –і–Є—Б–њ—Г—В–µ –Љ–љ–Њ–≥–Є–µ –њ—А–Є–Љ–µ—А—Л –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –±—Л–ї–Є –≤–Ј—П—В—Л –њ–Њ –і–∞–љ–љ—Л–Љ –Ј–∞—А—Г–±–µ–ґ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–µ–є, —З—В–Њ –Ј–∞—В—А—Г–і–љ—П–ї–Њ –Њ–±—А–∞—В–љ—Г—О —Б–≤—П–Ј—М —Б –∞–≤—В–Њ—А–∞–Љ–Є, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, —Б—В–∞–ї–Њ –Њ–±–Є–і–љ–Њ –Ј–∞ –±—Л–≤—И—Г—О 1/6 —З–∞—Б—В—М —Б—Г—И–Є, –љ–∞ –Ї–Њ—В–Њ—А–Њ–є –≤—Б–µ –Љ—Л –њ—А–Њ–ґ–Є–≤–∞–µ–Љ, –∞ –љ–µ–Ї–Њ—В–Њ—А—Л–µ –њ—А–Њ–ґ–Є–≤–∞–ї–Є —А–∞–љ—М—И–µ.–Э–µ—Г–ґ–µ–ї–Є –≤ –†–Њ—Б—Б–Є–Є –њ–µ—А–µ–≤–µ–ї–Є—Б—М –Я—А–ґ–µ–≤–∞–ї—М—Б–Ї–Є–µ, –∞ —Б–∞–Љ–∞ —Н–≤–Њ–ї—О—Ж–Є—П –Њ–±–Є–і–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –Њ–±–Њ—И–ї–∞ –љ–∞—Б –Є —В—Г—В —Б—В–Њ—А–Њ–љ–Њ–є? –Ъ–∞–Ї –≤—Л—П—Б–љ–Є–ї–Њ—Б—М, –≤ —В–∞–Ї–Њ–Љ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–µ–љ–Є–Є —П –Њ—И–Є–±–∞–ї—Б—П –Є –і–Њ–≤–Њ–ї—М–љ–Њ —Б–µ—А—М—С–Ј–љ–Њ.
–Ш—В–∞–Ї, –µ—Й—С —А–∞–Ј –љ–∞–њ–Њ–Љ–љ—О –Є—Б—Е–Њ–і–љ—Л–µ —Г—Б–ї–Њ–≤–Є—П, –Ї–Њ—В–Њ—А—Л–Љ–Є –њ—А–Є—И–ї–Њ—Б—М —А—Г–Ї–Њ–≤–Њ–і—Б—В–≤–Њ–≤–∞—В—М—Б—П –њ—А–Є –њ–Њ–Є—Б–Ї–∞—Е : –∞—В–µ–Є—Б—В–Њ–≤ –њ—А–Њ—Б–Є–ї–Є –њ—А–Є–≤–µ—Б—В–Є —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ—Л–є –њ—А–Є–Љ–µ—А –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –≤ –ґ–Є–≤–Њ–є –њ—А–Є—А–Њ–і–µ, —Б –∞–љ–∞–ї–Є–Ј–Њ–Љ –Є —А–∞–Ј—К—П—Б–љ–µ–љ–Є–µ–Љ –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є—Е, –Є—Б—В–Њ—А–Є—З–µ—Б–Ї–Є—Е, –Ї–ї–Є–Љ–∞—В–Є—З–µ—Б–Ї–Є—Е –Є –њ—А–Њ—З–Є—Е –њ—А–Є—З–Є–љ —Б—Г—Й–љ–Њ—Б—В–Є –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П (–Є –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–љ–Њ–є —Н—В–Є–Љ –≤–Є–і–Њ–≤–Њ–є –Є–Ј–Њ–ї—П—Ж–Є–Є), –∞ –і–ї—П –њ—А–Є–Љ–µ—А–∞ –њ—А–Њ—Б–Є–ї–Є –≤–Ј—П—В—М –і–Њ–≤–Њ–ї—М–љ–Њ —Б–ї–Њ–ґ–љ–Њ–µ –њ–Њ –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є–Є (—Г—Б—В—А–Њ–є—Б—В–≤—Г) –ґ–Є–≤–Њ–µ —Б—Г—Й–µ—Б—В–≤–Њ.–Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –њ—А–Њ—Б–Є–ї–Є –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞—В—М –Є –Њ–±—К—П—Б–љ–Є—В—М вАУ –Ї–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ –љ–∞–Ї–Њ–њ–ї–µ–љ–Є–µ –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є—Е —А–∞–Ј–ї–Є—З–Є–є –≤ –Ї–∞—А–Є–Њ—В–Є–њ–∞—Е –њ–Њ —Е–Њ–і—Г –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –њ—А–Є–≤–Њ–і–Є—В –Ї –≤–Є–і–Њ–≤–Њ–є –Є–Ј–Њ–ї—П—Ж–Є–Є.
–Т –Ї–∞—З–µ—Б—В–≤–µ –њ—А–Є–Љ–µ—А–∞, –њ–Њ—Б–ї–µ –і–ї–Є—В–µ–ї—М–љ—Л—Е –њ–Њ–Є—Б–Ї–Њ–≤ –≤ —Б–µ—В–Є, –Љ–љ–µ –Њ—З–µ–љ—М –≥–ї—П–љ—Г–ї–∞—Б—М —Б—В–∞—В—М—П –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤–∞ –Є –Я.–Ь.–С–Њ—А–Њ–і–Є–љ–∞: вАЬ–Ю–±—Л–Ї–љ–Њ–≤–µ–љ–љ–∞—П –±—Г—А–Њ–Ј—Г–±–Ї–∞: –•—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–є –њ–Њ—А—В—А–µ—В –љ–∞ —Д–Њ–љ–µ –ї–µ–і–љ–Є–Ї–Њ–≤вАЭ. –ѓ –±—Л–ї–Њ —Б–њ–Є—Б–∞–ї—Б—П (–≤ –Ї–Њ–љ—Ж–µ 2001–≥.) —Б –≥–ї–∞–≤–љ—Л–Љ —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –њ–Њ —Н–≤–Њ–ї—О—Ж–Є–Є –≤ вАЬ–Ш–Ј–±–µ-–Ф–µ–±–∞—В–љ–µвАЭ - –Ь.–Ы–Њ–±–∞–љ–Њ–≤—Л–Љ. –Ґ–Њ—В –Ј–∞–≤–Њ—А—З–∞–ї –≤ –Њ—В–≤–µ—В, —З—В–Њ –і–∞–љ–љ–∞—П —Б—В–∞—В—М—П –љ–µ –њ–Њ–ї–љ–∞—П, –љ–µ —Е–≤–∞—В–∞–µ—В —А–Є—Б—Г–љ–Ї–Њ–≤, —В–∞–±–ї–Є—Ж –Є –Њ—Б—В–∞–ї—М–љ—Л—Е –Є–ї–ї—О—Б—В—А–∞—Ж–Є–є, –Ї–Њ–µ-—З—В–Њ –љ–µ —П—Б–љ–Њ –њ–Њ —В–µ–Ї—Б—В—Г, –њ–Њ—Н—В–Њ–Љ—Г –љ–µ–Є–Ј–±–µ–ґ–љ—Л –≤–Њ–њ—А–Њ—Б—Л –Њ–њ–њ–Њ–љ–µ–љ—В–Њ–≤, –љ–∞ –Ї–Њ—В–Њ—А—Л–µ –Љ—Л –љ–µ –Њ—В–≤–µ—В–Є–Љ.–Ю–і–Є–љ –Є–Ј –≥–ї–∞–≤–љ—Л–є –Њ–њ–њ–Њ–љ–µ–љ—В–Њ–≤ –Њ—В –≤–µ—А—Г—О—Й–Є—Е –≤ вАЬ–Ш–Ј–±–µ-–Ф–µ–±–∞—В–љ–µвАЭ –Р-—Б–∞–є—В–∞ - –Ч—Л—А—П–љ–Є–љ –љ–∞—З–∞–ї –Љ–µ–ґ–і—Г —В–µ–Љ —Б–µ—В–Њ–≤–∞—В—М, —З—В–Њ –і–Њ—Б—В–Њ–є–љ–Њ–≥–Њ –њ—А–Є–Љ–µ—А–∞ —Г —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–Є—Б—В–Њ–≤ –њ—А–Њ—Б—В–Њ –љ–µ—В. –Я—А–Є—И–ї–Њ—Б—М –њ—Г—В—С–Љ –њ–µ—А–µ–≥–Њ–≤–Њ—А —Г—Б–ї–Њ–≤–Є—В—М—Б—П, —З—В–Њ –Ї—А–∞–є–љ–Є–Љ —Б—А–Њ–Ї–Њ–Љ –њ—А–µ–і–Њ—Б—В–∞–≤–ї–µ–љ–Є—П –њ—А–Є–Љ–µ—А–∞ —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–Є—Б—В–∞–Љ–Є –±—Г–і–µ—В –љ–∞—З–∞–ї–Њ –Т–µ–ї–Є–Ї–Њ–≥–Њ –њ–Њ—Б—В–∞ –≤ 2002 –≥.(–і–Њ 5 –Љ–∞—А—В–∞).–Я–Њ–ї—Г—З–Є–≤ –≤—А–µ–Љ—П –љ–∞ –њ–Њ–Є—Б–Ї–Є –њ—А–Є–Љ–µ—А–∞, —П –љ–∞–њ–Є—Б–∞–ї –њ–Є—Б—М–Љ–Њ –љ–∞ —Б–∞–є—В www.nature.ru —Б –њ—А–Њ—Б—М–±–Њ–є –і–∞—В—М –Ї–Њ–љ—В–∞–Ї—В–љ—Л–µ –∞–і—А–µ—Б–∞ –∞–≤—В–Њ—А–Њ–≤ —Б—В–∞—В—М–Є –Њ –±—Г—А–Њ–Ј—Г–±–Ї–∞—Е. –Ь–љ–µ –Њ—В–≤–µ—В–Є–ї–Є, —З—В–Њ –µ—Б—В—М —Б–∞–є—В, –≥–і–µ —П –Љ–Њ–≥—Г –њ–Њ–њ—А–Њ–±–Њ–≤–∞—В—М –њ–Њ–Є—Б–Ї–∞—В—М –∞–≤—В–Њ—А–Њ–≤ –≤ —Б–њ–Є—Б–Ї–µ —Б–Њ—В—А—Г–і–љ–Є–Ї–Њ–≤ , –≥–і–µ –і–µ–є—Б—В–≤–Є—В–µ–ї—М–љ–Њ –љ–∞—И—С–ї—Б—П —Н–ї–µ–Ї—В—А–Њ–љ–љ. –∞–і—А–µ—Б –Њ–і–љ–Њ–≥–Њ –Є–Ј –∞–≤—В–Њ—А–Њ–≤ —Б—В–∞—В—М–Є вАУ –Я.–Ь.–С–Њ—А–Њ–і–Є–љ–∞.–Т –Њ—В–≤–µ—В –љ–∞ –Љ–Њ—С –њ–Є—Б—М–Љ–Њ, –Я.–Ь.–С–Њ—А–Њ–і–Є–љ –і–∞–ї –∞–і—А–µ—Б –і—А—Г–≥–Њ–≥–Њ —Б–Њ–∞–≤—В–Њ—А–∞ вАУ –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤–∞.–ѓ –Њ—В–њ—А–∞–≤–Є–ї –њ–Є—Б—М–Љ–Њ –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤—Г –Є –Њ–љ –Љ–љ–µ –ї—О–±–µ–Ј–љ–Њ –Њ—В–≤–µ—В–Є–ї –љ–∞ —З–Є—Б—В–µ–є—И–µ–Љ –∞–љ–≥–ї–Є–є—Б–Ї–Њ–Љ, —З—В–Њ —Б–µ–є—З–∞—Б –љ–∞—Е–Њ–і–Є—В—Б—П –Ј–∞ –≥—А–∞–љ–Є—Ж–µ–є, –љ–µ –Є–Љ–µ–µ—В –љ–∞ –Ї–Њ–Љ–њ—М—О—В–µ—А–µ –Ї–Є—А–Є–ї–ї–Є—Ж—Л –Є –Љ—Л —Б –љ–Є–Љ –Љ–Њ–ґ–µ–Љ —В–Њ–ї—М–Ї–Њ "—Б–њ–Є–Ї–∞—В—М".–ѓ –Њ—З–µ–љ—М —Б–Њ–Љ–љ–µ–≤–∞–ї—Б—П –Ј–∞ —Б–≤–Њ–є –∞–љ–≥–ї–Є–є—Б–Ї–Є–є –≤ —З–∞—Б—В–Є —В–Њ—З–љ—Л—Е –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Б–њ–µ—Ж—В–µ—А–Љ–Є–љ–Њ–≤ (—В.–Ї. –љ–µ —П–≤–ї—П—О—Б—М –±–Є–Њ–ї–Њ–≥–Њ–Љ), –љ–Њ –љ–µ —Г–і–µ—А–ґ–∞–ї—Б—П –Є –љ–∞ —А–∞–і–Њ—Б—В—П—Е, –Њ—В —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–љ–Њ–≥–Њ –Ї–Њ–љ—В–∞–Ї—В–∞ —Б –Њ–і–љ–Є–Љ –Є–Ј –∞–≤—В–Њ—А–Њ–≤ –њ—Г–±–ї–Є–Ї–∞—Ж–Є–Є, –љ–∞–њ—А–∞–≤–Є–ї –і–ї–Є–љ–љ—О—Й–Є–є —Б–њ–Є—Б–Њ–Ї –≤–Њ–њ—А–Њ—Б–Њ–≤ –њ–Њ –±—Г—А–Њ–Ј—Г–±–Ї–∞–Љ –љ–∞ –љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е –ї–Є—Б—В–∞—Е.. –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤ —Г–і–Є–≤–Є—В–µ–ї—М–љ–Њ –±—Л—Б—В—А–Њ –Љ–љ–µ –Њ—В–≤–µ—В–Є–ї, —З—В–Њ –≤—Б—С –њ–Њ–љ—П–ї –Є –њ—А–Є—Б–ї–∞–ї –≤ –Њ—В–≤–µ—В —Ж–µ–ї—Г—О —Б—В–∞—В—М—О –љ–∞ –∞–љ–≥–ї–Є–є—Б–Ї–Њ–Љ —П–Ј—Л–Ї–µ –Є–Ј –љ–∞—Г—З–љ–Њ–≥–Њ –ґ—Г—А–љ–∞–ї–∞ вАЬRussian Journal of GeneticsвАЭ, Vol 37, No 4, 2001, pp.351-357 .(–°–Љ. –њ—А–Є–ї–Њ–ґ–µ–љ–Є–µ 1). –Э–Є—З–µ–≥–Њ –і—А—Г–≥–Њ–≥–Њ –≤ —В–Њ—В –Љ–Њ–Љ–µ–љ—В –Њ –±—Г—А–Њ–Ј—Г–±–Ї–∞—Е —Г –љ–µ–≥–Њ –њ–Њ–і —А—Г–Ї–Њ–є( —З—В–Њ–±—Л –Њ—В–њ—А–∞–≤–Є—В—М –Љ–љ–µ) –љ–µ –Њ–Ї–∞–Ј–∞–ї–Њ—Б—М.–Ф–∞–љ–љ–∞—П –њ—Г–±–ї–Є–Ї–∞—Ж–Є—П, –њ–Њ –µ–≥–Њ –Љ–љ–µ–љ–Є—О, –і–Њ–ї–ґ–љ–∞ –±—Л–ї–∞ –Њ—В–≤–µ—В–Є—В—М –љ–∞ 99% –Љ–Њ–Є—Е –≤–Њ–њ—А–Њ—Б–Њ–≤. –Т–ї–∞–і–µ—О—Й–Є–µ –∞–љ–≥–ї–Є–є—Б–Ї–Є–Љ –Љ–Њ–≥—Г—В –Њ–Ј–љ–∞–Ї–Њ–Љ–Є—В—М—Б—П —Б–Њ —Б—В–∞—В—М—С–є , –∞ —З–Є—В–∞—В–µ–ї–Є —Б –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ–є –њ–Њ–і–≥–Њ—В–Њ–≤–Ї–Њ–є –Љ–Њ–≥—Г—В –њ—А–Њ–њ—Г—Б—В–Є—В—М –њ—А–Є–ї–Њ–ґ–µ–љ–Є–µ 1, —В.–Ї. –љ–Є–ґ–µ –±—Г–і–µ—В –і–∞–љ —А—Г—Б—Б–Ї–Њ—П–Ј—Л—З–љ—Л–є –≤–∞—А–Є–∞–љ—В —Н—В–Њ–є —Б—В–∞—В—М–Є.
–°–≥–Њ—А—П—З–∞ —П —Г–ґ–µ —Б–Њ–±–Є—А–∞–ї—Б—П –Ј–∞—Б–µ—Б—В—М –Ј–∞ –њ–Њ–ї–љ—Л–є –њ–µ—А–µ–≤–Њ–і –≤—Б–µ–є —Б—В–∞—В—М–Є –љ–∞ —А—Г—Б—Б–Ї–Є–є, –Ї–Њ–≥–і–∞ –≤—Л—Б–Љ–Њ—В—А–µ–ї –≤ –Ј–∞–≥–Њ–ї–Њ–≤–Ї–µ –∞–љ–≥–ї–Є–є—Б–Ї–Њ–є –≤–µ—А—Б–Є–Є , —З—В–Њ —Н—В–Њ –њ–µ—А–µ–≤–Њ–і –Є–Ј —А—Г—Б—Б–Ї–Њ–≥–Њ –ґ—Г—А–љ–∞–ї–∞ вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ, #4, vol.37, 2001–≥. –Т—Б—С –Њ—Б—В–∞–ї—М–љ–Њ–µ –±—Л–ї–Њ –і–µ–ї–Њ–Љ –≤—А–µ–Љ–µ–љ–Є.–ѓ –њ–Њ–Љ—З–∞–ї—Б—П –≤ –†–У–С (–±—Л–≤—И—Г—О –Љ–Њ—Б–Ї–Њ–≤—Б–Ї—Г—О –±–Є–±–ї–Є–Њ—В–µ–Ї—Г –Є–Љ.–Ы–µ–љ–Є–љ–∞) –Є —З–µ—А–µ–Ј 20 –Љ–Є–љ—Г—В –Њ–ґ–Є–і–∞–љ–Є—П, –њ–µ—А–µ–і–Њ –Љ–љ–Њ–є –ї–µ–ґ–∞–ї –Є—Б–Ї–Њ–Љ—Л–є –ґ—Г—А–љ–∞–ї —Б –Њ—А–Є–≥–Є–љ–∞–ї–Њ–Љ —Б—В–∞—В—М–Є –љ–∞ —А—Г—Б—Б–Ї–Њ–Љ —П–Ј—Л–Ї–µ.–Х—Й—С 20 –Љ–Є–љ—Г—В + 20 —А—Г–±–ї–µ–є –Ј–∞—В—А–∞—В –Є —Г –Љ–µ–љ—П –љ–∞ —А—Г–Ї–∞—Е –±—Л–ї–∞ –Ї—Б–µ—А–Њ–Ї–Њ–њ–Є—П –і–∞–љ–љ–Њ–є —Б—В–∞—В—М–Є.–Т –Є—В–Њ–≥–µ —В–∞–Ї–Њ–≥–Њ –Ј–∞–љ–Є–Љ–∞—В–µ–ї—М–љ–Њ–≥–Њ –њ–Њ–Є—Б–Ї–∞, —Г–і–∞–ї–Њ—Б—М –њ–Њ–ї—Г—З–Є—В—М –њ–µ—А–≤–Њ–Є—Б—В–Њ—З–љ–Є–Ї –≤ –Њ—А–Є–≥–Є–љ–∞–ї–µ.–Ъ–∞–Ї –љ–∞–Ј–ї–Њ, –Є–Ј –і–≤—Г—Е –њ—А–Є–≤—Л—З–љ—Л—Е –Љ–љ–µ –Љ–µ—Б—В –Њ—В—Б–Ї–∞–љ–Є—А–Њ–≤–∞—В—М —Б—В–∞—В—М—О (–і–ї—П –Р-—Б–∞–є—В–∞) - –Њ–±–∞ –Њ–Ї–∞–Ј–∞–ї–Є—Б—М –≤—А–µ–Љ–µ–љ–љ–Њ –љ–µ–і–Њ—Б—В—Г–њ–љ—Л–Љ–Є, –≤–≤–Є–і—Г –њ–Њ–≤—Б–µ–Љ–µ—Б—В–љ–Њ —А–∞–Ј–≤–µ—А–љ—Г–≤—И–µ–є—Б—П –і–µ–Ї–∞–і—Л –њ—А–µ–і/–њ–Њ—Б–ї–µ –љ–Њ–≤–Њ–≥–Њ–і–љ–Є—Е —Б–і–≤–Є–≥–∞–љ–Є–є —Б—В–Њ–ї–Њ–≤ –Є —А–∞—Б—Б–ї–∞–±–ї–µ–љ–Є—П —В—А—Г–і–Њ–≤—Л—Е –Ї–Њ–ї–ї–µ–Ї—В–Є–≤–Њ–≤. –Ґ–µ–Ї—Б—В–∞ –±—Л–ї–Њ –љ–∞ 2-3 —З–∞—Б–∞ –љ–∞–±–Њ—А–∞, –∞ —А–Є—Б—Г–љ–Ї–Є –≤ –њ–µ—А–≤–Њ–Є—Б—В–Њ—З–љ–Є–Ї–µ –Є –∞–љ–≥–ї–Є–є—Б–Ї–Њ–Љ –≤–∞—А–Є–∞–љ—В–µ - –Њ–і–Є–љ–∞–Ї–Њ–≤—Л–Љ–Є.–ѓ —А–∞—Б—Б—Г–і–Є–ї, —З—В–Њ –љ–µ –Ј–∞–Ј–Њ—А–љ–Њ –љ–∞–±—А–∞—В—М —В–µ–Ї—Б—В —Б–∞–Љ–Њ–Љ—Г –Є –њ—А–Є —Н—В–Њ–Љ –≤ –Њ—З–µ—А–µ–і–љ–Њ–є —А–∞–Ј –≤–љ–Є–Љ–∞—В–µ–ї—М–љ–Њ –њ–µ—А–µ—З–Є—В–∞—В—М –≤—Б—О —Б—В–∞—В—М—О, —В.–Ї. –Њ—В–≤–µ—В–Њ–≤ –љ–∞ –≤–Њ–њ—А–Њ—Б—Л –≤ –і–µ–±–∞—В–љ–µ –љ–µ –Є–Ј–±–µ–ґ–∞—В—М.–Я—А–µ–і–ї–∞–≥–∞—О –≤–љ–Є–Љ–∞–љ–Є—О –≥–Є–±—А–Є–і: –Љ–Њ–є –љ–∞–±–Њ—А —В–µ–Ї—Б—В–∞ + –Є–ї–ї—О—Б—В—А–∞—Ж–Є–Є –Є–Ј –∞–љ–≥–ї. –≤–∞—А–Є–∞–љ—В–∞ —Б—В–∞—В—М–Є, –Ї–Њ—В–Њ—А—Л–µ –љ–Є—З–µ–Љ –љ–µ –Њ—В–ї–Є—З–∞—О—В—Б—П –≤ —А—Г—Б—Б–Ї–Њ—П–Ј—Л—З–љ–Њ–є –≤–µ—А—Б–Є–Є. –Р —Б–Њ–≤—Б–µ–Љ –і–Њ—В–Њ—И–љ—Л–Љ —А–µ–Ї–Њ–Љ–µ–љ–і—Г—О –љ–∞–є—В–Є –Њ—А–Є–≥–Є–љ–∞–ї –ґ—Г—А–љ–∞–ї–∞ вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ N 4 ,vol.37, 2001–≥. –Є —Б–≤–µ—А–Є—В—М –њ—А–∞–≤–Є–ї—М–љ–Њ—Б—В—М –Љ–Њ–µ–≥–Њ –љ–∞–±–Њ—А–∞ —В–µ–Ї—Б—В–∞ —Б –њ–µ—А–≤–Њ–Є—Б—В–Њ—З–љ–Є–Ї–Њ–Љ.
–У–Х–Э–Х–Ґ–Ш–Ъ–Р, 2001, —В–Њ–Љ 37, N4, —Б 448-455
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Р–ѓ –≠–Т–Ю–Ы–Ѓ–¶–Ш–ѓ –Ю–С–Ђ–Ъ–Э–Ю–Т–Х–Э–Э–Ю–Щ –С–£–†–Ю–†–Ч–£–С–Ъ–ШSorex araneus L. –Т –Я–Ю–°–Ы–Х–Ы–Х–Ф–Э–Ш–Ъ–Ю–Т–Ю–Х –Т–†–Х–Ь–ѓ
–Э–Р –Ѓ–Ц–Э–Ю–Ь –£–†–Р–Ы–Х –Ш –Т –°–Ш–С–Ш–†–Ш.
2001–Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤(1), –Т.–Т, –Я–∞–љ–Њ–≤(2), –Ґ.–Ѓ.–Ы–∞–і—Л–≥–Є–љ–∞(1), –Ь.–Э.–С–Њ—З–Ї–∞—А–µ–≤(1), –Ь.–Ш. –†–Њ–і–Є–Њ–љ–Њ–≤–∞(1), –Я.–Ь.–С–Њ—А–Њ–і–Є–љ(1, 3).
(1)-–Ш–љ—Б—В–Є—В—Г—В —Ж–Є—В–Њ–ї–Њ–≥–Є–Є –Є –≥–µ–љ–µ—В–Є–Ї–Є –°–Є–±–Є—А—Б–Ї–Њ–≥–Њ –Њ—В–і–µ–ї–µ–љ–Є—П –†–Р–Э, –Э–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї 630090;
—Д–∞–Ї—Б:(3832) 33-12-78; e-mail: polyakov@bionet.nsc.ru
(2) –Ш–љ—Б—В–Є—В—Г—В —Б–Є—Б—В–µ–Љ–∞—В–Є–Ї–Є –Є —Н–Ї–Њ–ї–Њ–≥–Є–Є –ґ–Є–≤–Њ—В–љ—Л—Е –°–Є–±–Є—А—Б–Ї–Њ–≥–Њ –Њ—В–і–µ–ї–µ–љ–Є—П –†–Р–Э,
–Э–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї 630091; —Д–∞–Ї—Б (3832) 35-65-58
(3)-–Э–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Є–є –≥–Њ—Б—Г–і–∞—А—Б—В–≤–µ–љ–љ—Л–є —Г–љ–Є–≤–µ—А—Б–Є—В–µ—В, –Э–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї 630090
–Я–Њ—Б—В—Г–њ–Є–ї–∞ –≤ —А–µ–і–∞–Ї—Ж–Є—О 28.08.2000.
–Я–Њ–і–≤–Њ–і—П—В—Б—П –Є—В–Њ–≥–Є —Б–µ—А–Є–Є —А–∞–±–Њ—В –њ–Њ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ–Њ–є –≥–µ–Њ–≥—А–∞—Д–Є–Є –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є (Sorex araneus L)
–≤ –°–Є–±–Є—А–Є –Є –љ–∞ –Ѓ–ґ–љ–Њ–Љ –£—А–∞–ї–µ.–•—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ —А–∞—Б—Л, –љ–∞—Б–µ–ї—П—О—Й–Є–µ —О–≥ –£—А–∞–ї–∞ –Є –Ч–∞–њ–∞–і–љ–Њ-–°–Є–±–Є—А—Б–Ї—Г—О —А–∞–≤–љ–Є–љ—Г, –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ —Б–Љ–µ–љ—П—О—В –і—А—Г–≥ –і—А—Г–≥–∞ –≤ —И–Є—А–Њ—В–љ–Њ–Љ –љ–∞–њ—А–∞–≤–ї–µ–љ–Є–Є. –Ъ–∞—А–Є–Њ—В–Є–њ—Л –ї—О–±—Л—Е –і–≤—Г—Е —Б–Њ—Б–µ–і–љ–Є—Е —А–∞—Б –љ–∞ —Н—В–Њ–є —В–µ—А—А–Є—В–Њ—А–Є–Є –Њ—В–ї–Є—З–∞—О—В—Б—П –і—А—Г–≥ –Њ—В –і—А—Г–≥–∞ –µ–і–Є–љ—Б—В–≤–µ–љ–љ–Њ–є –њ–Њ–ї–љ–Њ–њ–ї–µ—З–µ–≤–Њ–є —А–µ—Ж–Є–њ—А–Њ–Ї–љ–Њ–є —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є–µ–є.–Т –Ї–Њ–Љ–њ–ї–µ–Ї—Б–µ —А–∞—Б –Т–Њ—Б—В–Њ—З–љ–Њ–є –°–Є–±–Є—А–Є –Є –Р–ї—В–∞—П —Б–Љ–µ–ґ–љ—Л–µ —А–∞—Б—Л –Њ—В–ї–Є—З–∞—О—В—Б—П –≥–ї–∞–≤–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –њ–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤—Г –Є —Б–Њ—Б—В–∞–≤—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ.–Р–љ–∞–ї–Є–Ј —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е —А–∞—Б –≤ —Б–Њ—З–µ—В–∞–љ–Є–Є —Б –і–∞–љ–љ—Л–Љ–Є –њ–Њ –њ–∞–ї–µ–Њ—Н–Ї–Њ–ї–Њ–≥–Є–Є –ї–µ–і–љ–Є–Ї–Њ–≤–Њ–≥–Њ –Є –њ–Њ—Б–ї–µ–ї–µ–і–љ–Є–Ї–Њ–≤–Њ–≥–Њ –њ–µ—А–Є–Њ–і–Њ–≤ –њ–Њ–Ј–≤–Њ–ї—П–µ—В —А–µ–Ї–Њ–љ—Б—В—А—Г–Є—А–Њ–≤–∞—В—М –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В—М —Б—В–∞–љ–Њ–≤–ї–µ–љ–Є—П —Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л –≤–Є–і–∞ S. araneus.
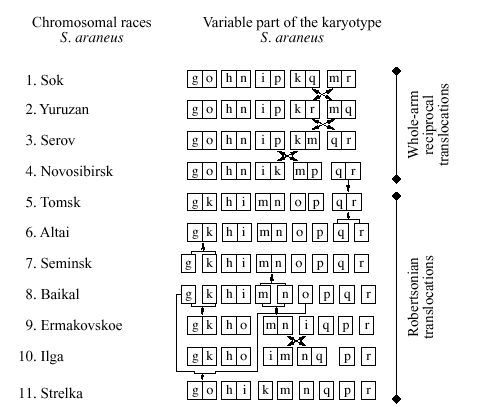
–Ю—Б–Њ–±–µ–љ–љ–Њ—Б—В—М—О –Ї–∞—А–Є–Њ—В–Є–њ–Є—З–µ—Б–Ї–Њ–є –Є–Ј–Љ–µ–љ—З–Є–≤–Њ—Б—В–Є –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є (Sorex araneus L) —П–≤–ї—П–µ—В—Б—П –њ–Њ–ї–Є–Љ–Њ—А—Д–Є–Ј–Љ –њ–Њ —А–Њ–±–µ—А—В—Б–Њ–љ–Њ–≤—Б–Ї–Є–Љ —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є—П–Љ: –њ—А–Є –љ–µ–Є–Ј–Љ–µ–љ–љ–Њ–Љ —З–Є—Б–ї–µ –∞—Г—В–Њ—Б–Њ–Љ–љ—Л—Е –њ–ї–µ—З(FNa=36), –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –∞—Г—В–Њ—Б–Њ–Љ –≤–∞—А—М–Є—А—Г–µ—В—Б—П –Њ—В 2Na=18 –і–Њ 2Na=30.–°–Њ–≥–ї–∞—Б–љ–Њ —Б—В–∞–љ–і–∞—А—В–љ–Њ–є –љ–Њ–Љ–µ–љ–Ї–ї–∞—В—Г—А–µ, —Г —Н—В–Њ–≥–Њ –≤–Є–і–∞ –Ї–∞–ґ–і–Њ–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ–Њ–µ –њ–ї–µ—З–Њ –Њ–±–Њ–Ј–љ–∞—З–∞–µ—В—Б—П –±—Г–Ї–≤–Њ–є –ї–∞—В–Є–љ—Б–Ї–Њ–≥–Њ –∞–ї—Д–∞–≤–Є—В–∞[1] .–Р—Г—В–Њ—Б–Њ–Љ—Л af, bc, –Є tu - –Є–љ–≤–∞—А–Є–∞–љ—В–љ—Л–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Є.–Ю—Б—В–∞–ї—М–љ—Л–µ 12 –∞—Г—В–Њ—Б–Њ–Љ–љ—Л—Е –њ–ї–µ—З - g, h, i, j, k, l, m, n, o, p, q, r - –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞—О—В—Б—П –ї–Є–±–Њ –≤ –≤–Є–і–µ –∞–Ї—А–Њ—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ, –ї–Є–±–Њ –≤ —А–∞–Ј–ї–Є—З–љ—Л—Е –њ–∞—А–љ—Л—Е –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є—П—Е –і—А—Г–≥ —Б –і—А—Г–≥–Њ–Љ –≤ —Б–Њ—Б—В–∞–≤–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ.[2,3] .–Ъ–∞—А–Є–Њ—В–Є–њ –Ї–∞–ґ–і–Њ–є –њ–Њ–њ—Г–ї—П—Ж–Є–Є –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –Њ–њ–Є—Б—Л–≤–∞–µ—В—Б—П –Њ–њ—А–µ–і–µ–ї–µ–љ–љ—Л–Љ —Б–Њ—З–µ—В–∞–љ–Є–µ–Љ –≤–∞—А–Є–∞–±–µ–ї—М–љ—Л—Е –њ–ї–µ—З –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ.–У—А—Г–њ–њ—Л –њ–Њ–њ—Г–ї—П—Ж–Є–є —Б –Њ–і–Є–љ–∞–Ї–Њ–≤—Л–Љ–Є –Ї–∞—А–Є–Њ—В–Є–њ–∞–Љ–Є, –љ–∞—Б–µ–ї—П—О—Й–Є–µ –µ–і–Є–љ—Г—О —В–µ—А—А–Є—В–Њ—А–Є—О, –±—Л–ї–Є –љ–∞–Ј–≤–∞–љ—Л —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–Љ–Є —А–∞—Б–∞–Љ–Є.[3] .–Ъ –љ–∞—Б—В–Њ—П—Й–µ–Љ—Г –≤—А–µ–Љ–µ–љ–Є –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –≤–µ—Б—М –∞—А–Є–∞–ї –≤–Є–і–∞ Sorex araneus L –Є–Ј—Г—З–µ–љ, –Є –±—Л–ї–Њ –њ–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ –µ–≥–Њ –Ї–∞—А–Є–Њ—В–Є–њ–Є—З–µ—Б–Ї–∞—П —Б—В—А—Г–Ї—В—Г—А–∞ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В —Б–Њ–±–Њ–є –Љ–Њ–Ј–∞–Є–Ї—Г –њ–∞—А–∞–њ–∞—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б.–С–Њ–ї–µ–µ 50 —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б –Њ–њ–Є—Б–∞–љ–Њ –љ–∞ —Б–µ–≥–Њ–і–љ—П—И–љ–Є–є –і–µ–љ—М.[2,3] .
–Ъ–∞—А–Є–Њ—В–Є–њ—Л –њ–Њ–і–∞–≤–ї—П—О—Й–µ–≥–Њ –±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ —А–∞—Б (9 –Є–Ј 11), –љ–∞—Б–µ–ї—П—О—Й–Є—Е –°–Ї–∞–љ–і–Є–љ–∞–≤—Б–Ї–Є–є –њ–Њ–ї—Г–Њ—Б—В—А–Њ–≤ –њ–Њ–ї–љ–Њ—Б—В—М—О —Б–Њ—Б—В–Њ—П—В –Є–Ј –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ.–†–∞—Б—Л –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ —Б–Љ–µ–љ—П—О—В –і—А—Г–≥ –і—А—Г–≥–∞, –њ—А–Є—З—С–Љ –Ї–∞–ґ–і–∞—П –њ–Њ—Б–ї–µ–і—Г—О—Й–∞—П —А–∞—Б–∞ –Њ—В–ї–Є—З–∞–µ—В—Б—П –Њ—В –њ—А–µ–і—Л–і—Г—Й–µ–є –µ–і–Є–љ—Б—В–≤–µ–љ–љ–Њ–є –њ–Њ–ї–љ–Њ–њ–ї–µ—З–µ–≤–Њ–є —А–µ—Ж–Є–њ—А–Њ–Ї–љ–Њ–є —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є–µ–є.[4,5] .–Т –Њ—Б—В–∞–ї—М–љ–Њ–є —З–∞—Б—В–Є –Х–≤—А–Њ–њ—Л –ї–Є—И—М 4 –Є–Ј 26 —А–∞—Б –Є–Љ–µ—О—В –њ–Њ–ї–љ–Њ—Б—В—М—О –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–є –Ї–∞—А–Є–Њ—В–Є–њ, –њ—А–Є —Н—В–Њ–Љ –Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ–µ —З–Є—Б–ї–Њ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ —Б–Њ–і–µ—А–ґ–Є—В—Б—П –≤ –Ї–∞—А–Є–Њ—В–Є–њ–∞—Е —А–∞—Б, –Њ–±–Є—В–∞—О—Й–Є—Е –љ–∞ —О–ґ–љ–Њ–є –Њ–Ї—А–∞–Є–љ–µ –µ–≤—А–Њ–њ–µ–є—Б–Ї–Њ–≥–є —З–∞—Б—В–Є –∞—А–µ–∞–ї–∞ S araneus (–®–≤–µ–є—Ж–∞—А–Є—П, –Ґ—Г—А—Ж–Є—П).[2,3}.
–У–Є–њ–Њ—В–µ–Ј—Л –Њ–± —Н–≤–Њ–ї—О—Ж–Є–Є S araneus —Б–≤—П–Ј—Л–≤–∞—О—В –њ—А–Њ–Є—Б—Е–Њ–ґ–і–µ–љ–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б —Б –≥–ї–Њ–±–∞–ї—М–љ—Л–Љ–Є –Є–Ј–Љ–µ–љ–µ–љ–Є—П–Љ–Є –Ї–ї–Є–Љ–∞—В–∞ –≤ –њ–µ—А–Є–Њ–і –њ–Њ—Б–ї–µ–і–љ–µ–≥–Њ –Њ–ї–µ–і–µ–љ–µ–љ–Є—П [6-8] .–Т –Њ—Б–љ–Њ–≤–µ —Н—В–Є—Е –≥–Є–њ–Њ—В–µ–Ј –ї–µ–ґ–Є—В –њ—А–µ–і–њ–Њ–ї–Њ–ґ–µ–љ–Є–µ –Њ —В–Њ–Љ, —З—В–Њ –≤ –Є—Б—Е–Њ–і–љ–Њ–Љ –њ—А–µ–і–Ї–Њ–≤–Њ–Љ –Ї–∞—А–Є–Њ—В–Є–њ–µ –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –≤—Б–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л –≤–∞—А–Є–∞–±–µ–ї—М–љ–Њ–є —З–∞—Б—В–Є –±—Л–ї–Є –∞–Ї—А–Њ—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–Љ–Є.–Т–Њ –≤—А–µ–Љ—П –Њ–ї–µ–і–µ–љ–µ–љ–Є—П –∞—А–µ–∞–ї –≤–Є–і–∞ S araneus –±—Л–ї —Д—А–∞–≥–Љ–µ–љ—В–Є—А–Њ–≤–∞–љ.–Т–Њ–Ј–љ–Є–Ї–ї–Њ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –љ–µ–±–Њ–ї—М—И–Є—Е –Є–Ј–Њ–ї–Є—А–Њ–≤–∞–љ–љ—Л—Е –і—А—Г–≥ –Њ—В –і—А—Г–≥–∞ –њ–Њ–њ—Г–ї—П—Ж–Є–є, –≤ –Ї–Њ—В–Њ—А—Л—Е –њ—А–Њ–Є—Б—Е–Њ–і–Є–ї–Є —А–Њ–±–µ—А—В—Б–Њ–љ–Њ–≤—Б–Ї–Є–µ —Б–ї–Є—П–љ–Є—П –Є —Д–Є–Ї—Б–∞—Ж–Є—П –≤–љ–Њ–≤—М –Њ–±—А–∞–Ј—Г—О—Й–Є—Е—Б—П –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤.–Я–Њ–Ј–ґ–µ, —Б –њ–Њ—В–µ–њ–ї–µ–љ–Є–µ–Љ, –∞—А–µ–∞–ї—Л —Н—В–Є—Е –њ–Њ–њ—Г–ї—П—Ж–Є–є —А–∞—Б—И–Є—А—П–ї–Є—Б—М –Є —Б–Љ—Л–Ї–∞–ї–Є—Б—М; –≤ –Љ–µ—Б—В–∞—Е –Є—Е –Ї–Њ–љ—В–∞–Ї—В–Њ–≤ –≤–Њ–Ј–љ–Є–Ї–∞–ї–Є —Г–Ј–Ї–Є–µ –Ј–Њ–љ—Л –Є–љ—В–µ—А–≥—А–∞–і–∞—Ж–Є–Є [3, 7-9] . –Э–∞ —В–µ—А—А–Є—В–Њ—А–Є–Є –°–Ї–∞–љ–і–Є–љ–∞–≤–Є–Є —А–∞—Б—Л —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ - –Њ–і–љ–∞ –Ј–∞ –Њ–і–љ–Њ–є, –њ–Њ –Љ–µ—А–µ –Ј–∞—Б–µ–ї–µ–љ–Є—П –±—Г—А–Њ–Ј—Г–±–Ї–∞–Љ–Є –њ–Њ–ї—Г–Њ—Б—В—А–Њ–≤–∞ [4,5,10] .–Т —В–Њ –≤—А–µ–Љ—П, –Ї–Њ–≥–і–∞ —А–∞–Ј—А–∞–±–∞—В—Л–≤–∞–ї–Є—Б—М —Н—В–Є –≥–Є–њ–Њ—В–µ–Ј—Л, –і–∞–љ–љ—Л–µ –њ–Њ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ–Њ–є –і–Є—Д—Д–µ—А–µ–љ—Ж–Є–∞—Ж–Є–Є S. araneus –љ–∞ —В–µ—А—А–Є—В–Њ—А–Є–Є –°–Є–±–Є—А–Є –Є –£—А–∞–ї–∞ –±—Л–ї–Є –≤–µ—Б—М–Љ–∞ –љ–µ–Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л –Є, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –љ–µ –њ—А–Є–љ–Є–Љ–∞–ї–Є—Б—М –≤–Њ –≤–љ–Є–Љ–∞–љ–Є–µ –≤ —Д–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є—Е —А–µ–Ї–Њ–љ—Б—В—А—Г–Ї—Ж–Є—П—Е.
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Р–ѓ –≠–Т–Ю–Ы–Ѓ–¶–Ш–ѓ –Ю–С–Ђ–Ъ–Э–Ю–Т–Х–Э–Э–Ю–Щ –С–£–†–Ю–Ч–£–С–Ъ–Ш
–†–Є—Б.1 –§–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–µ –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј–Є —О–ґ–љ–Њ—Г—А–∞–ї—М—Б–Ї–Є—Е –Є —Б–Є–±–Є—А—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є.
–Ф–Њ 1995 –≥. –њ–Њ –Њ—В–і–µ–ї—М–љ—Л–Љ –љ–∞—Е–Њ–і–Ї–∞–Љ –±—Л–ї–Є –Њ–њ–Є—Б–∞–љ—Л —З–µ—В—Л—А–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ —А–∞—Б—Л —Н—В–Њ–≥–Њ –≤–Є–і–∞ –≤ –°–Є–±–Є—А–Є.[11-14] .–†–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ —Н—В–Є—Е —А–∞—Б –Њ—Б—В–∞–≤–∞–ї–Њ—Б—М –љ–µ—П—Б–љ—Л–Љ.
–Т 1995-1997 –≥–≥. –Љ—Л –њ—А–Њ–≤–µ–ї–Є —А—П–і —Н–Ї—Б–њ–µ–і–Є—Ж–Є–є, –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ –Ї–Њ—В–Њ—А—Л—Е –≤ –°–Є–±–Є—А–Є –Є –љ–∞ –£—А–∞–ї–µ –±—Л–ї–Њ –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ 5 —А–∞–љ–µ–µ –љ–µ –Є–Ј–≤–µ—Б—В–љ—Л—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б.–Ф–ї—П –љ–µ–Ї–Њ—В–Њ—А—Л—Е –љ–Њ–≤—Л—Е –Є –њ—А–µ–ґ–і–µ –Њ–њ–Є—Б–∞–љ–љ—Л—Е —А–∞—Б –±—Л–ї–Є –њ—А–Є–±–ї–Є–Ј–Є—В–µ–ї—М–љ–Њ –Њ—З–µ—А—З–µ–љ—Л –≥—А–∞–љ–Є—Ж—Л –Є—Е –≥–µ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П.–Ф–∞–љ–љ–∞—П —Б—В–∞—В—М—П —Б—Г–Љ–Љ–Є—А—Г–µ—В –≤–µ—Б—М –љ–∞—И –Љ–∞—В–µ—А–Є–∞–ї –њ–Њ –≥–µ–Њ–≥—А–∞—Д–Є–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –≤–∞—А–Є–∞–љ—В–Њ–≤ –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є.–Т—Б–µ –Њ–±–љ–∞—А—Г–ґ–µ–љ–љ—Л–µ –љ–∞–Љ–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ —А–∞—Б—Л –Њ–њ–Є—Б–∞–љ—Л –≤ —Б–µ—А–Є–Є –њ—Г–±–ї–Є–Ї–∞—Ж–Є–є –≤–Љ–µ—Б—В–µ —Б –њ—А–Є–Љ–µ–љ—П–≤—И–Є–Љ–Є—Б—П –і–ї—П –∞–љ–∞–ї–Є–Ј–∞ –Љ–µ—В–Њ–і–∞–Љ–Є.[15-21] .
–°–Њ–≤–Њ–Ї—Г–њ–љ–Њ—Б—В—М –Є–Љ–µ—О—Й–Є—Е—Б—П —Г –љ–∞—Б –і–∞–љ–љ—Л—Е –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П –Ј–і–µ—Б—М –і–ї—П –Њ–±—Б—Г–ґ–і–µ–љ–Є—П —Н–≤–Њ–ї—О—Ж–Є–Є S. araneus –≤ –°–Є–±–Є—А–Є –Є –љ–∞ –£—А–∞–ї–µ –Є, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, –і–ї—П —А–∞–Ј–≤–Є—В–Є—П –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є—П –Њ –њ—Г—В—П—Е —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—П —Н—В–Њ–≥–Њ –≤–Є–і–∞ –≤ —Ж–µ–ї–Њ–Љ.
–Ю–°–Ю–С–Х–Э–Э–Ю–°–Ґ–Ш –Ъ–Р–†–Ш–Ю–Ґ–Ш–Я–Ю–Т –Ѓ–Ц–Э–Ю–£–†–Р–Ы–ђ–°–Ъ–Ш–• –Ш –°–Ш–С–Ш–†–°–Ъ–Ш–•
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Ђ–• –†–Р–° S. araneus –Ш –Ш–• –†–Р–°–Я–†–Ю–°–Ґ–†–Р–Э–Х–Э–Ш–Х.
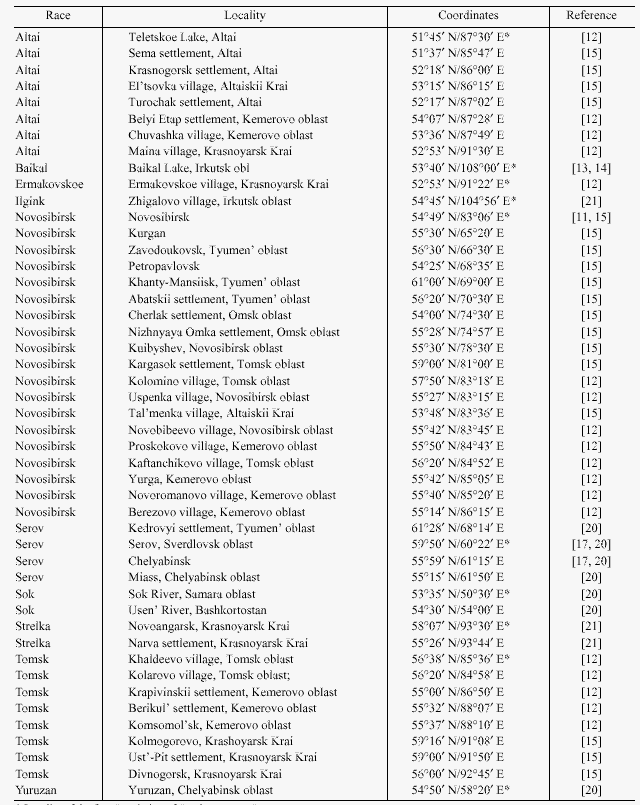
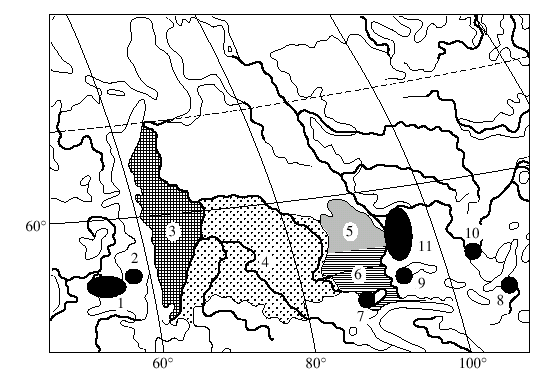
–Т–Њ—Б—В–Њ—З–љ—Л–є –Ї—А–∞–є –∞—А–µ–∞–ї–∞ –≤–Є–і–∞ S. araneus –љ–∞—Б–µ–ї–µ–љ –Ї–∞–Ї –Љ–Є–љ–Є–Љ—Г–Љ –і–≤—Г–Љ—П —А–∞—Б–∞–Љ–Є : –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є [13,14] –Є –Њ–±–љ–∞—А—Г–ґ–µ–љ–љ–Њ–є –љ–∞–Љ–Є –Є–ї–≥–Є–љ—Б–Ї–Њ–є [21] (—А–Є—Б. 1,2; —В–∞–±–ї–Є—Ж–∞).–С–∞–є–Ї–∞–ї—М—Б–Ї–∞—П —А–∞—Б–∞ —Б–∞–Љ–∞—П "–∞–Ї—А–Њ—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–∞—П" –≤ –°–Є–±–Є—А–Є: –≤—Б–µ–≥–Њ 4 –Є–Ј 12 –њ–ї–µ—З –≤–∞—А–Є–∞–±–µ–ї—М–љ–Њ–є —З–∞—Б—В–Є –µ—С –Ї–∞—А–Є–Њ—В–Є–њ–∞ –Њ–±—К–µ–і–Є–љ–µ–љ—Л –≤ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Є hi –Є jl.–Ы–Њ–Ї–∞–ї—М–љ–Њ—Б—В—М –Њ–њ–Є—Б–∞–љ–Є—П –Є–ї–≥–Є–љ—Б–Ї–Њ–є —А–∞—Б—Л –љ–∞—Е–Њ–і–Є—В—Б—П –њ—А–Є–±–ї–Є–Ј–Є—В–µ–ї—М–љ–Њ –≤ 300 –Ї–Љ —Б–µ–≤–µ—А–љ–µ–µ –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є.–Т –Њ—В–ї–Є—З–Є–µ –Њ—В –њ–Њ—Б–ї–µ–і–љ–µ–є –Є–ї–≥–Є–љ—Б–Ї–∞—П —А–∞—Б–∞ –љ–∞—Б—Л—Й–µ–љ–∞ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–Љ–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–∞–Љ–Є - –Ј–і–µ—Б—М —В–Њ–ї—М–Ї–Њ –і–≤–∞ –Љ–µ–ї–Ї–Є—Е –њ–ї–µ—З–∞ p –Є r —Б–Њ—Е—А–∞–љ–Є–ї–Є—Б—М –Ї–∞–Ї –∞–Ї—А–Њ—Ж–µ–љ—В—А–Є–Ї–Є.
–Ъ–∞—А–Є–Њ—В–Є–њ –µ—А–Љ–∞–Ї–Њ–≤—Б–Ї–Њ–є —А–∞—Б—Л, –Є–Ј–≤–µ—Б—В–љ–Њ–є –њ–Њ –µ–і–Є–љ—Б—В–≤–µ–љ–љ–Њ–Љ—Г –њ—Г–љ–Ї—В—Г (—А–Є—Б.1,2; —В–∞–±–ї–Є—Ж–∞), —В–∞–Ї–ґ–µ –љ–∞—Б—Л—Й–µ–љ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–Љ–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–∞–Љ–Є.[12] .–Т —Н—В–Њ–Љ –Њ—В–љ–Њ—И–µ–љ–Є–Є , –Ї–∞–Ї, –≤–њ—А–Њ—З–µ–Љ –Є –њ–Њ —Б–Њ—Б—В–∞—В–∞–≤—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ , –µ—А–Љ–∞–Ї–Њ–≤—Б–Ї–∞—П –Є –Є–ї–≥–Є–љ—Б–Ї–Є–µ —А–∞—Б—Л –≤–µ—Б—М–Љ–∞ —Б—Е–Њ–і–љ—Л –Љ–µ–ґ–і—Г —Б–Њ–±–Њ–є, –Њ—В–ї–Є—З–∞—П—Б—М –ї–Є—И—М –Њ–і–љ–Њ–є –њ–Њ–ї–љ–Њ–њ–ї–µ—З–µ–≤–Њ–є —А–µ—Ж–Є–њ—А–Њ–Ї–љ–Њ–є —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є–µ–є.
–Ъ —Б–µ–≤–µ—А—Г –Њ—В –µ—А–Љ–∞–Ї–Њ–≤—Б–Ї–Њ–є –Љ—Л –Њ–±–љ–∞—А—Г–ґ–Є–ї–Є –љ–Њ–≤—Г—О —А–∞—Б—Г - —Б—В—А–µ–ї–Ї–Є–љ—Б–Ї—Г—О (—А–Є—Б. 1,2;—В–∞–±–ї–Є—Ж–∞)[21] .–Ю–љ–∞ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–∞ , –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –≤–і–Њ–ї—М –њ—А–∞–≤–Њ–≥–Њ –±–µ—А–µ–≥–∞ –Х–љ–Є—Б–µ—П –Ї —Б–µ–≤–µ—А—Г –Њ—В –µ–≥–Њ –њ—А–∞–≤–Њ–≥–Њ –њ—А–Є—В–Њ–Ї–∞ - —А. –Ь–∞–љ–∞.
–Ы–µ–≤—Л–є –±–µ—А–µ–≥ –Х–љ–Є—Б–µ—П –Ј–∞—Б–µ–ї–µ–љ –∞–ї—В–∞–є—Б–Ї–Њ–є –Є —В–Њ–Љ—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є S. araneus ( —В–∞–±–ї–Є—Ж–∞) [12, 13, 15, 18, 22] .–†–∞–Ј–ї–Є—З–Є–µ –≤ –Ї–∞—А–Є–Њ—В–Є–њ–∞—Е —Н—В–Є—Е —А–∞—Б –Њ–≥—А–∞–љ–Є—З–Є–≤–∞–µ—В—Б—П —В–µ–Љ, —З—В–Њ –≤ –∞–ї—В–∞–є—Б–Ї–Њ–є —А–∞—Б–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л q –Є r –≤—Б–µ–≥–і–∞ –∞–Ї—А–Њ—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–µ, –∞ –≤ —В–Њ–Љ—Б–Ї–Њ–є –Њ–љ–Є, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ –≤—Б—В—А–µ—З–∞—О—В—Б—П –≤ —Б–Њ—Б—В–∞–≤–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–∞ qr.–Ѓ–ґ–љ—Л–є —А—Г–±–µ–ґ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П –±—Г—А–Њ–Ј—Г–±–Њ–Ї , –Є–Љ–µ—О—Й–Є—Е –≤ —Б–≤–Њ—С–Љ –Ї–∞—А–Є–Њ—В–Є–њ–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї qr , –њ—А–Є–±–ї–Є–Ј–Є—В–µ–ї—М–љ–Њ —Б–Њ–≤–њ–∞–і–∞–µ—В —Б —Б 54 –≥—А–∞–і. —Б–µ–≤–µ—А–љ–Њ–є —И–Є—А–Њ—В—Л [15] .–Э–∞ —Н—В–Њ–Љ –Њ—Б–љ–Њ–≤–∞–љ–Є–Є –Љ–Њ–ґ–љ–Њ –њ–Њ–ї–Њ–≥–∞—В—М, —З—В–Њ —В–µ—А—А–Є—В–Њ—А–Є—П –Љ–µ–ґ–і—Г —А–µ–Ї–∞–Љ–Є –Х–љ–Є—Б–µ–є –Є –Ю–±—М –Ї —О–≥—Г –Њ—В 54-–є —И–Є—А–Њ—В—Л –Ј–∞—Б–µ–ї–µ–љ–∞ –∞–ї—В–∞–є—Б–Ї–Њ–є, –∞ –Ї —Б–µ–≤–µ—А—Г —В–Њ–Љ—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є.
–Я—Г–љ–Ї—В—Л, –≤ –Ї–Њ—В–Њ—А—Л—Е –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–Є —А–∞–Ј–ї–Є—З–љ—Л—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б S. Araneus
* –Ь–µ—Б—В–Њ –њ–µ—А–≤–Є—З–љ–Њ–≥–Њ –Њ–њ–Є—Б–∞–љ–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ–Њ–є —А–∞—Б—Л.
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Р–ѓ –≠–Т–Ю–Ы–Ѓ–¶–Ш–ѓ –Ю–С–Ђ–Ъ–Э–Ю–Т–Х–Э–Э–Ю–Щ –С–£–†–Ю–Ч–£–С–Ъ–Ш
–†–Є—Б.2.–†–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ —О–ґ–љ–Њ—Г—А–∞–ї—М—Б–Ї–Њ–є –Є —Б–Є–±–Є—А—Б–Ї–Њ–є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б
–Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –≤ –љ—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П. –†–∞—Б—Л –њ—А–Њ–љ—Г–Љ–µ—А–Њ–≤–∞–љ—Л –≤ —В–Њ–Љ –ґ–µ –њ–Њ—А—П–і–Ї–µ,
—З—В–Њ –Є –љ–∞ —А–Є—Б.1.
–Ч–∞–њ–∞–і–љ–µ–µ –∞–ї—В–∞–є—Б–Ї–∞—П –Є —В–Њ–Љ—Б–Ї–∞—П —А–∞—Б—Л –≥—А–∞–љ–Є—З–∞—В —Б –њ–Њ–ї–љ–Њ—Б—В—М—О "–Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Њ–є" –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є —А–∞—Б–Њ–є [11-13, 15, 16, 18, 22] .–Т–Њ—Б—В–Њ—З–љ–∞—П –≥—А–∞–љ–Є—Ж–∞ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є —А–∞—Б—Л –њ—А–Њ—Е–Њ–і–Є—В –њ–Њ –њ—А–∞–≤–Њ–Љ—Г –±–µ—А–µ–≥—Г —А. –Ю–±—М —Б —О–≥–∞ –љ–∞ —Б–µ–≤–µ—А, –Є –Є –њ–Њ—Б–ї–µ 55 –≥—А–∞–і. —Б.—И. –Њ—В–Ї–ї–Њ–љ—П—П—Б—М –љ–∞ –≤–Њ—Б—В–Њ–Ї –і–Њ—Б—В–Є–≥–∞–µ—В —А. –Ґ–Њ–Љ—М [ 12, 13, 15, 18 22]. –Ъ–∞—А–Є–Њ—В–Є–њ—Л –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є —А–∞—Б—Л —Б –Њ–і–љ–Њ–є —Б—В–Њ—А–Њ–љ—Л –Є —В–Њ–Љ—Б–Ї–Њ-–∞–ї—В–∞–є—Б–Ї–Њ–є –≥—А—Г–њ–њ—Л —Б –і—А—Г–≥–Њ–є, —А–µ–Ј–Ї–Њ —А–∞–Ј–ї–Є—З–∞—О—В—Б—П –њ–Њ —Б–Њ—Б—В–∞–≤—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ.–≠—В–Њ, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г –Њ–і–Є–љ –Є–Ј –љ–µ–Љ–љ–Њ–≥–Є—Е –Є–Ј–≤–µ—Б—В–љ—Л—Е —Б–ї—Г—З–∞–µ–≤, –Ї–Њ–≥–і–∞ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –Ї–Њ–Љ–њ–Њ–Ј–Є—Ж–Є–Є —Б–Њ—Б–µ–і–љ–Є—Е —А–∞—Б —А–∞–Ј–ї–Є—З–∞—О—В—Б—П —Б—В–Њ–ї—М —Б–Є–ї—М–љ–Њ.
–Ъ –Ј–∞–њ–∞–і—Г –Њ—В —А. –Ю–±—М –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–∞—П —А–∞—Б–∞ –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –њ–Њ–ї–љ–Њ—Б—В—М—О –Ј–∞–љ–Є–Љ–∞–µ—В –Ч–∞–њ–∞–і–љ–Њ-–°–Є–±–Є—А—Б–Ї—Г—О —А–∞–≤–љ–Є–љ—Г.–У—А–∞–љ–Є—Ж–µ–є –Љ–µ–ґ–і—Г –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є –Є –≤–њ–µ—А–≤—Л–µ –Њ–њ–Є—Б–∞–љ–љ–Њ–є –љ–∞–Љ–Є —Б–µ—А–Њ–≤—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є —П–≤–ї—П—О—В—Б—П —А–µ–Ї–Є –Ґ–Њ–±–Њ–ї –Є –Ш—А—В—Л—И.[ 17, 19, 20]. –°–µ—А–Њ–≤—Б–Ї–∞—П —А–∞—Б–∞, —Б—Г–і—П –њ–Њ –љ–∞—И–µ–Љ—Г –Љ–∞—В–µ—А–Є–∞–ї—Г (—Б–Љ. —В–∞–±–ї–Є—Ж—Г), —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–∞ –њ–Њ –≤—Б–µ–є —В–µ—А—А–Є—В–Њ—А–Є–Є –≤–Њ—Б—В–Њ—З–љ—Л—Е –њ—А–µ–і–≥–Њ—А–Є–є –£—А–∞–ї–∞ –Є, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, –љ–∞ –њ—А–∞–≤–Њ–Љ –±–µ—А–µ–≥—Г —А–µ–Ї–Є –Ю–±—М, —Б–µ–≤–µ—А–љ–µ–µ –≤–њ–∞–і–µ–љ–Є—П –≤ –љ–µ—С —А. –Ш—А—В—Л—И.–Ъ–∞—А–Є–Њ—В–Є–њ —Б–µ—А–Њ–≤—Б–Ї–Њ–є —А–∞—Б—Л, –Ї–∞–Ї –Є –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є , —Б–Њ—Б—В–Њ–Є—В —В–Њ–ї—М–Ї–Њ –Є–Ј –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ.–Э–∞ –Ј–∞–њ–∞–і–љ—Л—Е —Б–Ї–ї–Њ–љ–∞—Е –£—А–∞–ї–∞ –Љ—Л –љ–∞—И–ї–Є –µ—Й—С –і–≤–µ –љ–Њ–≤—Л–µ —А–∞—Б—Л - —О—А—О–Ј–∞–љ—М—Б–Ї—Г—О –Є —Б–Њ–Ї—Б–Ї—Г—О.(—В–∞–±–ї–Є—Ж–∞) [20]., –≥—А–∞–љ–Є—Ж—Л —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П –Ї–Њ—В–Њ—А—Л—Е –њ—А–µ–і—Б—В–Њ–Є—В —Г—В–Њ—З–љ–Є—В—М.–Ъ–∞—А–Є–Њ—В–Є–њ—Л —Н—В–Є—Е –і–≤—Г—Е —А–∞—Б —В–∞–Ї–ґ–µ –њ–Њ–ї–љ–Њ—Б—В—М—О "–Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–µ".–Т–∞–ґ–љ–Њ –њ–Њ–і—З–µ—А–Ї–љ—Г—В—М, —З—В–Њ —Б–Њ–Ї—Б–Ї–∞—П, —О—А—О–Ј–∞–љ—М—Б–Ї–∞—П, —Б–µ—А–Њ–≤—Б–Ї–∞—П –Є –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–∞—П —А–∞—Б—Л, –њ–Њ–і–Њ–±–љ–Њ —Б–Ї–∞–љ–і–Є–љ–∞–≤—Б–Ї–Є–Љ, –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ —Б–Љ–µ–љ—П—О—В –і—А—Г–≥ –і—А—Г–≥–∞: –Ї–∞–ґ–і–∞—П –Є—Е –љ–Є—Е –Њ—В–ї–Є—З–∞–µ—В—Б—П –Њ—В —Б–Њ—Б–µ–і–љ–µ–є –њ–Њ –Њ–і–љ–Њ–є –њ–Њ–ї–љ–Њ–њ–ї–µ—З–µ–≤–Њ–є —А–µ—Ж–Є–њ—А–Њ–Ї–љ–Њ–є —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є–µ–є.(—Б–Љ. —А–Є—Б. 1).–Я—А–Є–Љ–µ—З–∞—В–µ–ї–µ–љ —В–Њ—В —Д–∞–Ї—В, —З—В–Њ —Г—А–∞–ї—М—Б–Ї–Є–µ —Б–Њ–Ї—Б–Ї–∞—П –Є —О—А—О–Ј–∞–љ—М—Б–Ї–∞—П —А–∞—Б—Л –Є–і–µ–љ—В–Є—З–љ—Л –њ–Њ —Б–Њ—Б—В–∞–≤—Г —Е—А–Њ–Љ–Њ—Б–Њ–Љ —А–∞—Б–∞–Љ –Ъ—Г–Љ–Њ –Є –Ш–ї—В–Њ–Љ–∞–љ—В—Б–Є —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ, –Ї–Њ—В–Њ—А—Л–µ –Ј–∞–Љ—Л–Ї–∞—О—В –љ–∞ –≤–Њ—Б—В–Њ–Ї–µ –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В—М —Б–Ї–∞–љ–і–Є–љ–∞–≤—Б–Ї–Є—Е —А–∞—Б.
–§–Ш–Ы–Ю–У–Х–Э–Х–Ґ–Ш–І–Х–°–Ъ–Ш–Х –°–Т–ѓ–Ч–Ш –Ь–Х–Ц–Ф–£ –Ш–°–°–Ы–Х–Ф–Ю–Т–Р–Э–Э–Ђ–Ь–Ш –•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Ђ–Ь–Ш –†–Р–°–Р–Ь–Ш –Ю–С–Ђ–Ъ–Э–Ю–Т–Х–Э–Э–Ю–Щ –С–£–†–Ю–Ч–£–С–Ъ–Ш.
–Я–Њ —Е–∞—А–∞–Ї—В–µ—А—Г –Ї–∞—А–Є–Њ—В–Є–њ–Є—З–µ—Б–Ї–Њ–є –Є–Ј–Љ–µ–љ—З–Є–≤–Њ—Б—В–Є –≤–љ—Г—В—А–Є –≤–Є–і–∞ S. araneus –≤ –°–Є–±–Є—А–Є –Є –љ–∞ –£—А–∞–ї–µ –Љ–Њ–ґ–љ–Њ –≤—Л–і–µ–ї–Є—В—М –і–≤–µ –љ–µ–Ј–љ–∞—З–Є—В–µ–ї—М–љ—Л–µ —Д–Є–ї–µ—В–Є—З–µ—Б–Ї–Є–µ –≤–µ—В–≤–Є.–Ю–і–љ–∞ –Є–Ј –љ–Є—Е –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–∞ —Б–Њ–Ї—Б–Ї–Њ–є, —О—А—О–Ј–∞–љ—М—Б–Ї–Њ–є, —Б–µ—А–Њ–≤—Б–Ї–Њ–є, –Є –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–Љ–Є —А–∞—Б–∞–Љ–Є, –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ —Б–Љ–µ–љ—П—О—Й–Є–Љ–Є –і—А—Г–≥ –і—А—Г–≥–∞ –≤ —И–Є—А–Њ—В–љ–Њ–Љ –љ–∞–њ—А–∞–≤–ї–µ–љ–Є–Є - –Њ—В –Ѓ–ґ–љ–Њ–≥–Њ –£—А–∞–ї–∞ –і–Њ –≤–Њ—Б—В–Њ—З–љ—Л—Е –њ—А–µ–і–µ–ї–Њ–≤ –Ч–∞–њ–∞–і–љ–Њ-–°–Є–±–Є—А—Б–Ї–Њ–є —А–∞–≤–љ–Є–љ—Л. –Ю —Е–∞—А–∞–Ї—В–µ—А–µ –Ї–∞—А–Є–Њ—В–Є–њ–Є—З–µ—Б–Ї–Њ–є –Є–Ј–Љ–µ–љ—З–Є–≤–Њ—Б—В–Є –≤ –і–∞–љ–љ–Њ–є –≥—А—Г–њ–њ–µ —А–∞—Б –≥–Њ–≤–Њ—А–Є–ї–Њ—Б—М –≤—Л—И–µ.–Ф—А—Г–≥–∞—П —Д–Є–ї–µ—В–Є—З–µ—Б–Ї–∞—П –≤–µ—В–≤—М –≤–Ї–ї—О—З–∞–µ—В –≤ —Б–µ–±—П –Ї–Њ–Љ–њ–ї–µ–Ї—Б —А–∞—Б –Т–Њ—Б—В–Њ—З–љ–Њ–є –°–Є–±–Є—А–Є –Є –Р–ї—В–∞—П. –≠—В–Њ—В –Ї–Њ–Љ–њ–ї–µ–Ї—Б –Њ—В–ї–Є—З–∞–µ—В—Б—П –Њ—В –њ—А–µ–і—Л–і—Г—Й–µ–≥–Њ –±–Њ–ї–µ–µ —Б–ї–Њ–ґ–љ–Њ–є –њ—А–Њ—Б—В—А–∞–љ—Б—В–≤–µ–љ–љ–Њ–є –Ї–Њ–љ—Д–Є–≥—Г—А–∞—Ж–Є–µ–є.–°–Љ–µ–ґ–љ—Л–µ —А–∞—Б—Л —А–∞—Б—Л –≤ –њ—А–µ–і–µ–ї–∞—Е –≤–Њ—Б—В–Њ—З–љ–Њ-—Б–Є–±–Є—А—Б–Ї–Њ–≥–Њ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞ —А–∞–Ј–ї–Є—З–∞—О—В—Б—П –≥–ї–∞–≤–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –њ–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤—Г –Є —Б–Њ—Б—В–∞–≤—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ, –Њ–±—А–∞–Ј–Њ–≤–∞–≤—И–Є—Е—Б—П –њ—Г—В—С–Љ —А–Њ–±–µ—А—В—Б–Њ–љ–Њ–≤—Б–Ї–Є—Е —Б–ї–Є—П–љ–Є–є.(—Б–Љ. —А–Є—Б—О. 1).–°–Њ–≥–ї–∞—Б–љ–Њ –љ–∞—И–Є–Љ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є—П–Љ, –Є—Б—Е–Њ–і–љ—Л–Љ –Ј–≤–µ–љ–Њ–Љ –≤ –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В–Є —А–∞—Б –≤–Њ—Б—В–Њ—З–љ–Њ-—Б–Є–±–Є—А—Б–Ї–Њ–є –≤–µ—В–≤–Є —П–≤–ї—П–µ—В—Б—П –±–∞–є–Ї–∞–ї—М—Б–Ї–∞—П —А–∞—Б–∞.(—А–Є—Б. 1).–Ю–љ–∞ –љ–∞—Б–µ–ї—П–µ—В —О–≥–Њ-–≤–Њ—Б—В–Њ—З–љ—Л–є –Ї—А–∞–є –∞—А–µ–∞–ї–∞ –≤–Є–і–∞, –∞ –≤–∞—А–Є–∞–±–µ–ї—М–љ–∞—П —З–∞—Б—В—М –µ—С –≥–∞–њ–ї–Њ–Є–і–љ–Њ–≥–Њ –Ї–∞—А–Є–Њ—В–Є–њ–∞ —Б–Њ–і–µ—А–ґ–Є—В –≤—Б–µ–≥–Њ –і–≤–∞ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–∞: hj –Є jl. –У–µ–љ–µ—В–Є—З–µ—Б–Ї–Є –љ–∞–Є–±–Њ–ї–µ–µ –±–ї–Є–Ј–Ї–Њ–є –Ї –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є —А–∞—Б–µ (–µ—Б–ї–Є —Б—Г–і–Є—В—М –њ–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ –≤ –Ї–∞—А–Є–Њ—В–Є–њ–µ) –Њ–Ї–∞–Ј—Л–≤–∞–µ—В—Б—П –∞–ї—В–∞–є—Б–Ї–∞—П —А–∞—Б–∞ ( gk hi jl); –Њ–љ–∞ –Љ–Њ–≥–ї–∞ –њ—А–Њ–Є–Ј–Њ–є—В–Є –Њ—В –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є —А–∞—Б—Л –њ—Г—В–µ–Љ –і–≤—Г—Е —Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Б–ї–Є—П–љ–Є–є.–Ч–і–µ—Б—М –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ –≤—Л–±–Њ—А–Ї–∞ –Є–Ј –њ—Г–љ–Ї—В–∞ –°–µ–Љ–∞ ( hi , jl, mn) –љ–∞ —О–ґ–љ–Њ–є –Њ–Ї—А–∞–Є–љ–µ –Ј–Њ–љ—Л —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П –∞–ї—В–∞–є—Б–Ї–Њ–є —А–∞—Б—Л [ 15, 21] –Љ–Њ–ґ–µ—В —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞—В—М—Б—П –Ї–∞–Ї –њ–µ—А–µ—Е–Њ–і–љ–∞—П —Д–Њ—А–Љ–∞ –Љ–µ–ґ–і—Г –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є –Є –∞–ї—В–∞–є—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є. –Ф–∞–ї–µ–µ –Ї —Б–µ–≤–µ—А—Г –Ј–∞ –∞–ї—В–∞–є—Б–Ї–Њ–є —А–∞—Б–Њ–є —Б–ї–µ–і—Г–µ—В —В–Њ–Љ—Б–Ї–∞—П( gk, hi, mn, qr), –Ї–Њ—В–Њ—А–∞—П –Њ—В–ї–Є—З–∞–µ—В—Б—П –Њ—В –њ—А–µ–і—Л–і—Г—Й–µ–є –љ–∞–ї–Є—З–Є–µ–Љ –≤ –Ї–∞—А–Є–Њ—В–Є–њ–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–∞ qr.–°—Г—Й–µ—Б—В–≤—Г–µ—В –і–≤–µ –≤–µ—А—Б–Є–Є –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –µ–≥–Њ –њ–Њ—П–≤–ї–µ–љ–Є—П –≤ –Ї–∞—А–Є–Њ—В–Є–њ–µ —В–Њ–Љ—Б–Ї–Њ–є —А–∞—Б—Л: 1) —Б–ї–Є—П–љ–Є–µ q+r -> qr –Є —Д–Є–Ї—Б–∞—Ж–Є—П –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ –≤ –Ї—А–∞–µ–≤—Л—Е —Б–µ–≤–µ—А–љ—Л—Е –њ–Њ–њ—Г–ї—П—Ж–Є—П—Е –∞–ї—В–∞–є—Б–Ї–Њ–є —А–∞—Б—Л –Є–ї–Є 2) –Є–љ—В—А–Њ–≥—А–µ—Б—Б–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л qr —Б –Ј–∞–њ–∞–і–∞, –Њ—В –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є —А–∞—Б—Л, —Г –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –Ї–Њ—В–Њ—А–Њ–є —Н—В–Њ—В –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї –Њ–±—Л—З–µ–љ.[15].–І—В–Њ–±—Л –Њ—В–і–∞—В—М –њ—А–µ–і–њ–Њ—З—В–µ–љ–Є–µ –Ї–∞–Ї–Њ–Љ—Г-–ї–Є–±–Њ –Є–Ј —Н—В–Є—Е –≤–∞—А–Є–∞–љ—В–Њ–≤, –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П.
–Ю—В –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є —А–∞—Б—Л. –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –Є —Б—В—А–µ–ї–Ї–Є–љ—Б–Ї–∞—П —А–∞—Б–∞, –Ї–Њ—В–Њ—А–∞—П —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–∞ –≤–і–Њ–ї—М –њ—А–∞–≤–Њ–≥–Њ –±–µ—А–µ–≥–∞ —А.–Х–љ–Є—Б–µ–є.–Ъ—А–Њ–Љ–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ hj –Є jl, —Е–∞—А–∞–Ї—В–µ—А–љ—Л—Е –і–ї—П –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є —А–∞—Б—Л, –Ї–∞—А–Є–Њ—В–Є–њ —Б—В—А–µ–ї–Ї–Є–љ—Б–Ї–Њ–є —А–∞—Б—Л —Б–Њ–і–µ—А–ґ–Є—В —В–∞–Ї–ґ–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї go (—А–Є—Б.1).
–Ф–ї—П —Г—В–Њ—З–љ–µ–љ–Є—П –њ—А–Њ–Є—Б—Е–Њ–ґ–і–µ–љ–Є—П –µ—А–Љ–∞–Ї–Њ–≤—Б–Ї–Њ–є –Є –Є–ї–≥–Є–љ—Б–Ї–Њ–є —А–∞—Б –љ—Г–ґ–љ—Л –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ—Л–µ –і–∞–љ–љ—Л–µ.–°–µ–є—З–∞—Б –љ–∞–Є–±–Њ–ї–µ–µ –≤–µ—А–Њ—П—В–љ—Л–Љ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В—Б—П –Є—Е —А–Њ–і—Б—В–≤–Њ —Б –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ-—В–Њ–Љ—Б–Ї–Њ–є –≤–µ—В–≤—М—О.–Х—А–Љ–∞–Ї–Њ–≤—Б–Ї–∞—П —А–∞—Б–∞ –Љ–Њ–≥–ї–∞ –њ—А–Њ–Є–Ј–Њ–є—В–Є –Є–Ј —Б–Њ—Б–µ–і–љ–µ–є –∞–ї—В–∞–є—Б–Ї–Њ–є –њ—Г—В—С–Љ –Њ–і–љ–Њ–є –њ–Њ–ї–љ–Њ–њ–ї–µ—З–µ–≤–Њ–є —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є–Є ( hi, o -> ho, i ), –∞ –Є–ї–≥–Є–љ—Б–Ї–∞—П - –Є–Ј –µ—А–Љ–∞–Ї–Њ–≤—Б–Ї–Њ–є –Ј–∞ —Б—З—С—В –Њ–і–љ–Њ–≥–Њ —Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Њ–≥–Њ —Б–ї–Є—П–љ–Є—П : i + q -> iq. –Т –Ї–∞—З–µ—Б—В–≤–µ –∞–ї—М—В–µ—А–љ–∞—В–Є–≤–љ–Њ–≥–Њ –≤–∞—А–Є–∞–љ—В–∞ –Љ–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–∞–≥–∞—В—М –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ–µ –њ—А–Њ–Є—Б—Е–Њ–ґ–і–µ–љ–Є–µ —Н—В–Є—Е —А–∞—Б –Њ—В –≥–Є–њ–Њ—В–µ—В–Є—З–µ—Б–Ї–Њ–є –њ—А–µ–і–Ї–Њ–≤–Њ–є —Д–Њ—А–Љ—Л —Б –Ї–∞—А–Є–Њ—В–Є–њ–Њ–Љ gk, ho, jl, i, m, n, p, q, r –Ј–∞ —Б—З—С—В –і–≤—Г—Е —А–Њ–±–µ—А—В—Б–Њ–љ–Њ–≤—Б–Ї–Є—Е —Б–ї–Є—П–љ–Є–є : i + m -> im, n + q -> nq –≤ —Б–ї—Г—З–∞–µ –Є–ї–≥–Є–љ—Б–Ї–Њ–є —А–∞—Б—Л , –Є –Њ–і–љ–Њ–≥–Њ —А–Њ–±–µ—А—В—Б–Њ–љ–Њ–≤—Б–Ї–Њ–≥–Њ —Б–ї–Є—П–љ–Є—П : m + n -> mn - –≤ —Б–ї—Г—З–∞–µ –µ—А–Љ–∞–Ї–Њ–≤—Б–Ї–Њ–є.
–Я—А–µ–і–ї–Њ–ґ–µ–љ–љ–∞—П –љ–∞–Љ–Є —Б—Е–µ–Љ–∞ —Н–≤–Њ–ї—О—Ж–Є–Є –Ї–∞—А–Є–Њ—В–Є–њ–Њ–≤ S. araneus –њ—Г—В–µ–Љ –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ–≥–Њ –љ–∞–Ї–Њ–њ–ї–µ–љ–Є—П –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ –Њ–±—К—П—Б–љ—П–µ—В —Е–∞—А–∞–Ї—В–µ—А–љ—Г—О –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В—М –≥–µ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞—Б–њ—А–µ–і–µ–ї–µ–љ–Є—П –Ї–∞—А–Є–Њ—В–Є–њ–Њ–≤ –≤ –∞–Ј–Є–∞—В—Б–Ї–Њ–є —З–∞—Б—В–Є –∞—А–µ–∞–ї–∞ —Н—В–Њ–≥–Њ –≤–Є–і–∞, –∞ –Є–Љ–µ–љ–љ–Њ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ —Б —О–≥–∞ –љ–∞ —Б–µ–≤–µ—А.
–Я–Р–Ы–Х–Ю–≠–Ъ–Ю–Ы–Ю–У–Ш–ѓ –Ш –†–Р–°–Ю–Ю–С–†–Р–Ч–Ю–Т–Р–Э–Ш–Х –£ –Ю–С–Ђ–Ъ–Э–Ю–Т–Х–Э–Э–Ю–Щ –С–£–†–Ю–Ч–£–С–Ъ–Ш.
–Я–Њ–Ј–і–љ–µ–є—И–Є–є –Љ–∞–Ї—А–Њ—Ж–Є–Ї–ї —З–µ—В–≤–µ—А—В–Є—З–љ–Њ–≥–Њ –њ–µ—А–Є–Њ–і–∞ –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М—О –Њ–Ї–Њ–ї–Њ 130 —В—Л—Б. –ї–µ—В –≤–Ї–ї—О—З–∞–µ—В –Љ–µ–ґ–ї–µ–і–љ–Є–Ї–Њ–≤—Г—О –Є –њ–Њ—Б–ї–µ–і–Њ–≤–∞–≤—И—Г—О –Ј–∞ –љ–Є–Љ –ї–µ–і–љ–Є–Ї–Њ–≤—Г—О —Н–њ–Њ—Е—Г.–Т–Њ –≤—А–µ–Љ—П –Љ–µ–ґ–ї–µ–ґ–љ–Є–Ї–Њ–≤—М—П, –Ї–Њ–≥–і–∞, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –Є —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї—Б—П S. araneus [23], –≤ –°–Є–±–Є—А–Є —Б—В—А—Г–Ї—В—Г—А–∞ –Ј–Њ–љ–∞–ї—М–љ–Њ—Б—В–Є –≤ —Ж–µ–ї–Њ–Љ –±—Л–ї–∞ –±–ї–Є–Ј–Ї–Њ–є –Ї —Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є.–Ь–Њ–ґ–љ–Њ –і—Г–Љ–∞—В—М, —З—В–Њ –Є —Б—В—А—Г–Ї—В—Г—А–∞ –Ї–∞—А–Є–Њ—В–Є–њ–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–Є—П –њ–Њ–њ—Г–ї—П—Ж–Є–є –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –њ—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ –љ–µ –Њ—В–ї–Є—З–∞–ї–∞—Б—М –Њ—В —В–Њ–є, —З—В–Њ –Љ—Л –љ–∞–±–ї—О–і–∞–µ–Љ —Б–µ–є—З–∞—Б –≤ —В–Њ–Љ —Б–Љ—Л—Б–ї–µ, —З—В–Њ –Є —В–Њ–≥–і–∞ —Б—Г—Й–µ—Б—В–≤–Њ–≤–∞–ї —И–Є—А–Њ—В–љ—Л–є –≥—А–∞–і–Є–µ–љ—В –њ–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤: —З–µ–Љ –і–∞–ї—М—И–µ –љ–∞ —Б–µ–≤–µ—А, —В–µ–Љ –Є—Е –±—Л–ї–Њ –±–Њ–ї—М—И–µ.
–Т —Н–њ–Њ—Е—Г –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ–≥–Њ –њ–Њ—Е–Њ–ї–Њ–і–∞–љ–Є—П 20 -18 —В—Л—Б. –ї–µ—В –љ–∞–Ј–∞–і –≤ –°–Є–±–Є—А–Є —Б–ї–Њ–ґ–Є–ї–∞—Б—М –Њ—З–µ–љ—М —Б—Г—А–Њ–≤–∞—П –Њ–±—Б—В–∞–љ–Њ–≤–Ї–∞.–≠—В–Њ –±—Л–ї–∞ —Н–њ–Њ—Е–∞ –Љ–Њ—Й–љ–Њ–є —Н–Ї—Б–њ–∞–љ—Б–Є–Є –Љ–љ–Њ–≥–Њ–ї–µ—В–љ–µ–є –Љ–µ—А–Ј–ї–Њ—В—Л, –Ї–Њ—В–Њ—А–∞—П –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –њ—А–Њ–і–≤–Є–љ—Г–≤—И–Є—Б—М –Ї —О–≥—Г, –≤—Л–Ј–≤–∞–ї–∞ –њ–Њ–ї–љ—Г—О –і–µ—Б—В—А—Г–Ї—Ж–Є—О –ї–µ—Б–љ–Њ–≥–Њ –њ–Њ—П—Б–∞.–Ш–Ј-–Ј–∞ —Б–њ–ї–Њ—И–љ–Њ–≥–Њ –ї–µ–і–љ–Є–Ї–Њ–≤–Њ–≥–Њ —Д—А–Њ–љ—В–∞ –љ–∞ —Б–µ–≤–µ—А–µ, –њ—А–µ–њ—П—В—Б—В–≤–Њ–≤–∞–≤—И–µ–≥–Њ —Б–≤–Њ–±–Њ–і–љ–Њ–Љ—Г —Б—В–Њ–Ї—Г –≤ –Ъ–∞—А—Б–Ї–Њ–µ –Љ–Њ—А–µ, –Ч–∞–њ–∞–і–љ–Њ-–°–Є–±–Є—А—Б–Ї–∞—П —А–∞–≤–љ–Є–љ–∞ –Њ–Ї–∞–Ј–∞–ї–∞—Б—М –≤–Њ –≤–ї–∞—Б—В–Є –≥–Є–≥–∞–љ—В—Б–Ї–Њ–≥–Њ –њ–Њ–і–њ—А—Г–і–љ–Њ–≥–Њ –Њ–Ј–µ—А–∞.–Ф—А–µ–љ–∞–ґ –Ј–і–µ—Б—М –Љ–Њ–≥ –Њ—Б—Г—Й–µ—Б—В–≤–ї—П—В—М—Б—П —В–Њ–ї—М–Ї–Њ —З–µ—А–µ–Ј –Ґ—Г—А–≥–∞–є—Б–Ї–Є–є –њ—А–Њ–≥–Є–±. –Т —В–Њ –≤—А–µ–Љ—П –≤ –°–Є–±–Є—А–Є —Б—Г—Й–µ—Б—В–≤–Њ–≤–∞–ї–∞ –ї–Є—И—М —В—А–Є –Ј–Њ–љ–∞–ї—М–љ—Л—Е —В–Є–њ–∞ —А–∞—Б—В–Є—В–µ–ї—М–љ–Њ—Б—В–Є - –њ–µ—А–Є–≥–ї—П—Ж–Є–∞–ї—М–љ–∞—П —В—Г–љ–і—А–Њ–≤–Њ—Б—В–µ–њ–љ–∞—П, —В—Г–љ–і—А–Њ–≤–∞—П –Є –ї–µ—Б–Њ—В—Г–љ–і—А–Њ–≤–∞—П. –Я–Њ—Б–ї–µ–і–љ—П—П —А–∞—Б–њ–Њ–ї–∞–≥–∞–ї–∞—Б—М —О–ґ–љ–µ–µ 56 –≥—А–∞–і. —Б.—И. –≤ –Я—А–Є—Г—А–∞–ї—М—Б–Ї–Њ–є –Є –Я—А–Є–µ–љ–Є—Б–µ–є—Б–Ї–Њ–є —З–∞—Б—В—П—Е —А–∞–≤–љ–Є–љ—Л, –∞ —В–∞–Ї–ґ–µ –њ–Њ —О–ґ–љ—Л–Љ –±–µ—А–µ–≥–∞–Љ –њ—А–Є–ї–µ–і–љ–Є–Ї–Њ–≤–Њ–≥–Њ –≤–Њ–і–Њ—С–Љ–∞.[24].–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —Д—А–∞–≥–Љ–µ–љ—В—Л –ї–µ—Б–∞ —Б–Њ—Е—А–∞–љ–Є–ї–Є—Б—М —В–Њ–≥–і–∞ –ї–Є—И—М –≤–і–Њ–ї—М —О–ґ–љ–Њ–є –Њ–Ї—А–∞–Є–љ—Л –Я–∞–ї–µ–∞—А–Ї—В–Є–Ї–Є.[25,26]. –•–Њ–ї–Љ—Л –Є –≥–Њ—А—Л –Ѓ–ґ–љ–Њ–≥–Њ –£—А–∞–ї–∞ –Є –Ѓ–ґ–љ–Њ–є –°–Є–±–Є—А–Є –Љ–Њ–≥–ї–Є –±—Л—В—М —А–µ—Д—Г–≥–Є—Г–Љ–∞–Љ–Є –і–ї—П –і—А–µ–≤–µ—Б–љ—Л—Е –≤–Є–і–Њ–≤ —А–∞—Б—В–µ–љ–Є–є [27]. –Є —Б–≤—П–Ј–∞–љ–љ—Л—Е —Б –љ–Є–Љ–Є –ґ–Є–≤–Њ—В–љ—Л—Е.[28]. –Ф–∞–љ–љ—Л–Љ–Є —А–µ—Д–Є–≥–Є—Г–Љ–∞–Љ–Є –Њ–≥—А–∞–љ–Є—З–Є–≤–∞–ї–Є—Б—М, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –Є –Њ–±–ї–∞—Б—В—М —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є.–Я–Њ –Љ–µ—А–µ —Б–Њ–Ї—А–∞—Й–µ–љ–Є—П —Б–µ–≤–µ—А–љ–Њ–є —З–∞—Б—В–Є –∞—А–µ–∞–ї–∞ –±—Г—А–Њ–Ј—Г–±–Њ–Ї –і–Њ–ї–ґ–љ–Њ –±—Л–ї–Њ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В—М –Є —Г–Љ–µ–љ—М—И–µ–љ–Є–µ –Є—Е –Ї–∞—А–Є–Њ—В–Є–њ–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–Є—П.–Э–∞—Б—Л—Й–µ–љ–љ—Л–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–∞–Љ–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ —А–∞—Б—Л, –Њ–±–Є—В–∞–≤—И–Є–µ –і–Њ –ї–µ–і–љ–Є–Ї–∞ –љ–∞ —Б–µ–≤–µ—А–µ –∞—А–µ–∞–ї–∞, —Б–Њ—Е—А–∞–љ—П–ї–Є—Б—М –ї–Є—И—М –љ–∞ –Ѓ–ґ–љ–Њ–Љ –£—А–∞–ї–µ, –∞ –љ–∞ –Ї—А–∞–є–љ–µ–Љ —О–≥–µ –∞—А–µ–∞–ї–∞ –≤ —А–µ—Д–Є–≥–Є—Г–Љ–∞—Е –°–∞—П–љ –Є –Р–ї—В–∞—П —Б–Њ—Е—А–∞–љ—П–ї–Є—Б—М –ї–Є—И—М –∞–Ї—А–Њ—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–µ —А–∞—Б—Л, –њ–Њ–і–Њ–±–љ—Л–µ –љ—Л–љ–µ—И–љ–µ–є –±–∞–є–Ї–∞–ї—М—Б–Ї–Њ–є —А–∞—Б–µ.–° –љ–∞—З–∞–ї–Њ–Љ –њ–Њ—В–µ–њ–ї–µ–љ–Є—П –Ї–ї–Є–Љ–∞—В–∞ –≤ –°–Є–±–Є—А–Є –ї–µ—Б–∞ —Б—В–∞–ї–Є —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ—П—В—М—Б—П –љ–∞ —Б–µ–≤–µ—А.–Т—Б–ї–µ–і –Ј–∞ –љ–Є–Љ–Є –љ–∞—З–∞–ї–Њ—Б—М —А–∞—Б—И–Є—А–µ–љ–Є–µ –∞—А–µ–∞–ї–∞ –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ—Л—Е –±—Г—А–Њ–Ј—Г–±–Њ–Ї.(—А–Є—Б.3).–Э–∞ —Б–µ–≤–µ—А–љ–Њ–Љ —Д—А–Њ–љ—В–µ –∞—А–µ–∞–ї–∞, –≤ –љ–∞–Є–±–Њ–ї–µ–µ —Н–Ї—Б—В—А–µ–Љ–∞–ї—М–љ—Л—Е –Є –љ–µ—Б—В–∞–±–Є–ї—М–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –Љ–Њ–≥–ї–Є –њ–µ—А–Є–Њ–і–Є—З–µ—Б–Ї–Є –≤–Њ–Ј–љ–Є–Ї–∞—В—М –Є–Ј–Њ–ї–Є—А–Њ–≤–∞–љ–љ—Л–µ –Љ–Є–Ї—А–Њ–њ–Њ–њ—Г–ї—П—Ж–Є–Є, –≤ –Ї–Њ—В–Њ—А—Л—Е –≤–µ—Б—М–Љ–∞ –≤–µ—А–Њ—П—В–љ–Њ –њ—А–Њ–Є—Б—Е–Њ–і–Є–ї–∞ —Д–Є–Ї—Б–∞—Ж–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е –њ–µ—А–µ—Б—В—А–Њ–µ–Ї.–Я–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ–µ –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є–µ –љ–Њ–≤—Л—Е –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ –Ј–∞ —Б—З—С—В —А–Њ–±–µ—А—В—Б–Њ–љ–Њ–≤—Б–Ї–Є—Е —Б–ї–Є—П–љ–Є–є –∞–Ї—А–Њ—Ж–µ–љ—В—А–Є–Ї–Њ–≤ –≤ –њ—А–Њ—Ж–µ—Б—Б–µ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П –±—Г—А–Њ–Ј—Г–±–Њ–Ї –љ–∞ —Б–µ–≤–µ—А –Є –Њ–њ—А–µ–і–µ–ї–Є–ї–Њ, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –Љ–µ—А–Є–і–Є–Њ–љ–∞–ї—М–љ—Л–є –≥—А–∞–і–Є–µ–љ—В –њ–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ –≤ –Ї–∞—А–Є–Њ—В–Є–њ–∞—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б –≤–Њ—Б—В–Њ—З–љ–Њ-—Б–Є–±–Є—А—Б–Ї–Њ–≥–Њ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞.
–•–†–Ю–Ь–Ю–°–Ю–Ь–Э–Р–ѓ –≠–Т–Ю–Ы–Ѓ–¶–Ш–ѓ –Ю–С–Ђ–Ъ–Э–Ю–Т–Х–Э–Э–Ю–Щ –С–£–†–Ю–Ч–£–С–Ъ–Ш.

–†–Є—Б.3 –Я—Г—В–Є —А–∞—Б—Б–µ–ї–µ–љ–Є—П –≤–Є–і–∞ Sorex araneus –≤ –њ–Њ—Б–ї–µ–ї–µ–і–љ–Є–Ї–Њ–≤–Њ–µ –≤—А–µ–Љ—П.
–°–Њ—Е—А–∞–љ–Є–≤—И–Є–µ—Б—П –≤ —А–µ—Д–Є–≥–Є—Г–Љ–∞—Е –Ѓ–ґ–љ–Њ–≥–Њ –£—А–∞–ї–∞ –ґ–Є–≤–Њ—В–љ—Л–µ, –≤ –Њ—В–ї–Є—З–Є–µ –Њ—В –Њ—В –±—Г—А–Њ–Ј—Г–±–Њ–Ї —О–ґ–љ—Л—Е —А–µ—Д–Є–≥–Є—Г–Љ–Њ–≤ –°–Є–±–Є—А–Є, –Њ–±–ї–∞–і–∞–ї–Є, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –њ–Њ–ї–љ–Њ—Б—В—М—О "–Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–Љ–Є" –Ї–∞—А–Є–Њ—В–Є–њ–∞–Љ–Є.–Я–Њ—Б–ї–µ –Њ—В—Б—В—Г–њ–ї–µ–љ–Є—П –ї–µ–і–љ–Є–Ї–∞ —Г—А–∞–ї—М—Б–Ї–Є–µ —Д–Њ—А–Љ—Л —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–Є–ї–Є—Б—М –љ–∞ –Ј–∞–њ–∞–і - —З–µ—А–µ–Ј —Б–µ–≤–µ—А –Т–Њ—Б—В–Њ—З–љ–Њ–є –Х–≤—А–Њ–њ—Л –≤ –§–Є–љ–ї—П–љ–і–Є—О –Є –і–∞–ї–µ–µ –≤ –®–≤–µ—Ж–Є—О –Є –љ–∞ –≤–Њ—Б—В–Њ–Ї, –і–Њ –≤–Њ—Б—В–Њ—З–љ–Њ–є –Њ–Ї—А–∞–Є–љ—Л –Ч–∞–њ–∞–і–љ–Њ-–°–Є–±–Є—А—Б–Ї–Њ–є —А–∞–≤–љ–Є–љ—Л.–Я—А–Є —Н—В–Њ–Љ —А–∞—Б–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є–µ —И–ї–Њ –њ–Њ –њ—Г—В–Є —Д–Є–Ї—Б–∞—Ж–Є–Є –њ–Њ–ї–љ–Њ–њ–ї–µ—З–µ–≤—Л—Е —А–µ—Ж–Є–њ—А–Њ–Ї–љ—Л—Е —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є–є –Љ–µ–ґ–і—Г –Љ–µ—В–∞—Ж–µ–љ—В—А–Є—З–µ—Б–Ї–Є–Љ–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ–∞–Љ–Є.
–Э–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–∞—П —А–∞—Б–∞, –≤–µ—А–Њ—П—В–љ–µ–µ –≤—Б–µ–≥–Њ, –њ—А–Њ–Є–Ј–Њ—И–ї–∞ –Њ—В —Б–µ—А–Њ–≤—Б–Ї–Њ–є —А–∞—Б—Л –Є —А–∞—Б—Б–µ–ї—П–ї–∞—Б—М –Њ—В –≤–Њ—Б—В–Њ—З–љ–Њ–є –Њ–Ї—А–∞–Є–љ—Л –£—А–∞–ї–∞ –љ–∞ –≤–Њ—Б—В–Њ–Ї, –Њ—Б–≤–∞–Є–≤–∞—П —В–µ—А—А–Є—В–Њ—А–Є—О, –Ї–Њ—В–Њ—А–∞—П –Њ—Б–≤–Њ–±–Њ–ґ–і–∞–ї–∞—Б—М –њ–Њ –Љ–µ—А–µ –≤—Л—Б—Л—Е–∞–љ–Є—П –Ч–∞–њ–∞–і–љ–Њ-–°–Є–±–Є—А—Б–Ї–Њ–≥–Њ –ї–µ–і–љ–Є–Ї–Њ–≤–Њ–≥–Њ –Њ–Ј–µ—А–∞.–Э–∞ –≤–Њ—Б—В–Њ–Ї–µ –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–∞—П —А–∞—Б–∞ —Б–Њ–Љ–Ї–љ—Г–ї–∞ —Б–≤–Њ–є –∞—А–µ–∞–ї —Б –∞—А–µ–∞–ї–Њ–Љ —В–Њ–Љ—Б–Ї–Њ-–∞–ї—В–∞–є—Б–Ї–Њ–≥–Њ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞ —А–∞—Б.–Э–∞ –≥—А–∞–љ–Є—Ж–µ –Љ–µ–ґ–і—Г –љ–Є–Љ–Є –≤–Њ–Ј–љ–Є–Ї–ї–∞ –≥–Є–±—А–Є–і–љ–∞—П –Ј–Њ–љ–∞, –Ї–Њ—В–Њ—А–∞—П —Б—Г—Й–µ—Б—В–≤—Г–µ—В –њ–Њ —Б–µ–є –і–µ–љ—М.[5, 22]. –≠—В–Є–Љ –Є –Њ–±—К—П—Б–љ—П–µ—В—Б—П —А–µ–Ј–Ї–Є–є –Ї–Њ–љ—В—А–∞—Б—В –њ–Њ —Б–Њ—Б—В–∞–≤—Г —Е—А–Њ–Љ–Њ—Б–Њ–Љ –Љ–µ–ґ–і—Г –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є —А–∞—Б–Њ–є –Є –∞–ї—В–∞–є—Б–Ї–Њ-—В–Њ–Љ—Б–Ї–Њ–є –≥—А—Г–њ–њ–Њ–є. –Ш—Е —А–∞–Ј–і–µ–ї—П—О—В –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —В—Л—Б—П—З –ї–µ—В –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ–є —Н–≤–Њ–ї—О—Ж–Є–Є.–Т –Љ–µ–є–Њ–Ј–µ —Г –≥–Є–±—А–Є–і–Њ–≤ –Љ–µ–ґ–і—Г –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є –Є —В–Њ–Љ—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є –і–Њ–ї–ґ–љ–∞ –Њ–±—А–∞–Ј–Њ–≤—Л–≤–∞—В—М—Б—П —Ж–µ–њ—М –Є–Ј 9 —Н–ї–µ–Љ–µ–љ—В–Њ–≤.–Я–Њ–і–Њ–±–љ—Л–µ –љ–∞—А—Г—И–µ–љ–Є—П, –Ї–∞–Ї –Є–Ј–≤–µ—Б—В–љ–Њ, –і–Њ–ї–ґ–љ—Л –њ—А–Є–≤–Њ–і–Є—В—М –Ї —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О –љ–µ—Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞–љ–љ—Л—Е –≥–∞–Љ–µ—В –Є, –Ї–∞–Ї —Б–ї–µ–і—Б—В–≤–Є–µ –Ї –њ–Њ–ї–љ–Њ–є —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є –Є–ї–Є –Ї —А–µ–Ј–Ї–Њ–Љ—Г —Б–љ–Є–ґ–µ–љ–Є—О –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В–Є –≥–Є–±—А–Є–і–љ—Л—Е –Њ—Б–Њ–±–µ–є [22,29].
–Ю–±–Њ–±—Й–∞—П –≤—Б–µ —Б–Ї–∞–Ј–∞–љ–љ–Њ–µ –≤—Л—И–µ, –Љ–Њ–ґ–љ–Њ —Б–і–µ–ї–∞—В—М
–Ј–∞–Ї–ї—О—З–µ–љ–Є–µ –Њ —В–Њ–Љ, —З—В–Њ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ —А–∞—Б—Л
–Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –≤ –°–Є–±–Є—А–Є —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М
–њ–Њ—Н—В–∞–њ–љ–Њ, –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ —Б –Њ—Б–≤–Њ–µ–љ–Є–µ–Љ –≤–Є–і–Њ–Љ
—В–µ—А—А–Є—В–Њ—А–Є–є, —Б—В–∞–љ–Њ–≤—П—Й–Є—Е—Б—П –і–Њ—Б—В—Г–њ–љ—Л–Љ–Є –њ–Њ –Љ–µ—А–µ
–њ–Њ—Б–ї–µ–ї–µ–і–љ–Є–Ї–Њ–≤–Њ–≥–Њ –њ–Њ—В–µ–њ–ї–µ–љ–Є—П –Ї–ї–Є–Љ–∞—В–∞.–Ь–Њ–ґ–љ–Њ
–њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ –Њ–±—Й–Є–µ –њ—А–Є–љ—Ж–Є–њ—Л —Н–≤–Њ–ї—О—Ж–Є–Є
–Ї–∞—А–Є–Њ—В–Є–њ–Њ–≤ –±—Г—А–Њ–Ј—Г–±–Ї–Є –±—Л–ї–Є —Б—Е–Њ–і–љ—Л–Љ–Є –≤ –Х–≤—А–Њ–њ–µ –Є –≤
–°–Є–±–Є—А–Є,–Ф–Њ–≤–Њ–ї—М–љ–Њ –Љ–Њ–љ–Њ—В–Њ–љ–љ—Л–є –ї–∞–љ–і—И–∞—Д—В –°–Є–±–Є—А–Є –Є
—Б—Г—А–Њ–≤—Л–є –Ї–ї–Є–Љ–∞—В –њ—А–Є–≤–µ–ї–Є –Ї —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О –Ј–і–µ—Б—М, –≤
–њ–Њ—Б–ї–µ–ї–µ–і–љ–Є–Ї–Њ–≤–Њ–µ –≤—А–µ–Љ—П –±–Њ–ї–µ–µ –њ—А–Њ—Б—В–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л
–≤–Є–і–∞ –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б
—В–∞–Ї–Њ–≤–Њ–є –≤ –Х–≤—А–Њ–њ–µ, —З—В–Њ —Б–і–µ–ї–∞–ї–Њ —Д–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є
–≤–Ј–∞–Є–Љ–Њ–Њ—В–љ–Њ—И–µ–љ–Є–µ—П –Љ–µ–ґ–і—Г —Б–Є–±–Є—А—Б–Ї–Є–Љ–Є —А–∞—Б–∞–Љ–Є –±–Њ–ї–µ–µ
–Њ—В—З–µ—В–ї–Є–≤—Л–Љ–Є.–Т –Х–≤—А–Њ–њ–µ –њ–Њ—Б–ї–µ–ї–µ–і–љ–Є–Ї–Њ–≤–∞—П –Љ–Є–≥—А–∞—Ж–Є—П
–≤–Є–і–∞ S. araneus –Є–Љ–µ–ї–∞, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –±–Њ–ї–µ–µ —Б–ї–Њ–ґ–љ—Л–є
—Е–∞—А–∞–Ї—В–µ—А, —З—В–Њ —Б–Ї–∞–Ј–∞–ї–Њ—Б—М –љ–∞ –Љ–µ–љ–µ–µ —Г–њ–Њ—А—П–і–Њ—З–µ–љ–љ–Њ–Љ
—А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–Є–Є –∞—А–µ–∞–ї–Њ–≤ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б
–Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –і—А—Г–≥ –і—А—Г–≥–∞.–Ю–і–љ–∞–Ї–Њ –Љ–µ—А–Є–і–Є–Њ–љ–∞–ї—М–љ—Л–є
–≥—А–∞–і–Є–µ–љ—В –њ–Њ —З–∞—Б—В–Њ—В–µ –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ –≤ –Ї–∞—А–Є–Њ—В–Є–њ–∞—Е
—Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б —Б—Е–Њ–і–µ–љ —Б —В–µ–Љ, —З—В–Њ –Љ—Л –љ–∞–±–ї—О–і–∞–µ–Љ –≤
–°–Є–±–Є—А–Є.–Ш—Б—Е–Њ–і—П –Є–Ј —Н—В–Њ–≥–Њ, –ї–Њ–≥–Є—З–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ
—Н–≤–Њ–ї—О—Ж–Є—П –Ї–∞—А–Є–Њ—В–Є–њ–Њ–≤ –±—Г—А–Њ–Ј—Г–±–Ї–Є –љ–∞—З–∞–ї–∞—Б—М –Ј–і–µ—Б—М,
–Ї–∞–Ї –Є –≤ –°–Є–±–Є—А–Є, —Б —Б —О–ґ–љ–Њ–є –Љ–∞—А–≥–Є–љ–∞–ї—М–љ–Њ–є –Њ–Ї—А–∞–Є–љ—Л –Є
—И–ї–∞ –њ–Њ –њ—Г—В–Є –љ–∞–Ї–Њ–њ–ї–µ–љ–Є—П –Љ–µ—В–∞—Ж–µ–љ—В—А–Є–Ї–Њ–≤ –њ–Њ –Љ–µ—А–µ
—А–∞—Б—И–Є—А–µ–љ–Є—П –∞—А–µ–∞–ї–∞ –љ–∞ —Б–µ–≤–µ—А.[30]. –°–Ї–∞–љ–і–Є–љ–∞–≤–Є—П –Є
–Т–Њ—Б—В–Њ—З–љ–∞—П –Х–≤—А–Њ–њ–∞ –Ј–∞—Б–µ–ї—П–ї–Є—Б—М —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–Љ–Є —А–∞—Б–∞–Љ–Є
—Г—А–∞–ї—М—Б–Ї–Њ–≥–Њ –њ—А–Њ–Є—Б—Е–Њ–ґ–і–µ–љ–Є—П.–≠—В–Є –і–≤–µ
—Д–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–µ –≤–µ—В–≤–Є - —О–ґ–љ–Њ–µ–≤—А–Њ–њ–µ–є—Б–Ї–∞—П –Є
—Г—А–∞–ї—М—Б–Ї–∞—П - –≤—Б—В—А–µ—В–Є–ї–Є—Б—М –≤ —А–∞–є–Њ–љ–µ —Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є
–Я–Њ–ї—М—И–Є.–Ш–Љ–µ–љ–љ–Њ —В–∞–Љ –Њ–њ–Є—Б–∞–љ–∞ –µ–і–Є–љ—Б—В–≤–µ–љ–љ–∞—П
–≥–Є–±—А–Є–і–љ–∞—П –Ј–Њ–љ–∞ (–Љ–µ–ґ–і—Г —А–∞—Б–∞–Љ–Є –Ф—А–љ—Е–Њ–ї–µ–Ї –Є
–С–µ–ї–Њ–≤–µ–ґ–∞), –Ї–Њ—В–Њ—А–∞—П –њ–Њ –Ї–Њ–љ—В—А–∞—Б—В–љ–Њ—Б—В–Є –Њ–±—А–∞–Ј–Њ–≤–∞–≤—И–Є—Е
–µ—С —А–∞—Б –Љ–Њ–ґ–µ—В —Б—А–∞–≤–љ–Є—В—М—Б—П —Б –≥–Є–±—А–Є–і–љ–Њ–є –Ј–Њ–љ–Њ–є –Љ–µ–ґ–і—Г
–љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є –Є –∞–ї—В–∞–є—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є –≤ –°–Є–±–Є—А–Є [ 31]
.
ACKNOWLEDGMENTS
We are grateful to A. Banashek, Ya. Zima, and
J. Searle for their invaluable help in collecting and ana-lyzing
the material and the reviewer of Genetika (Mos-cow)
for constructive comments.
This study was supported by the Russian Foun-dation
for Basic Research (grants no. 95-04-12698,
96-15-97738, 98-04-49734), the Ministry of Education
of the Russian Federation (grant no. 3H-220-98), and
INTAS (grant no. 93-1463).
C–°–Ђ–Ы–Ъ–Ш.
- Searle, J.B., Fedyk, S., Fredga, K., et al., Nomenclature for the Chromosomes of the Common Shrew (Sorex ara-neus), Memoires de la soci–єt–є vaudoise des sciences naturelles, 1991, vol. 19, pp. 13вАУ22.
- Zima, J., Fedyk, S., Fredga, K., et al., The List of the Chromosome Races of the Common Shrew (Sorex ara-neus), Hereditas (Lund, Swed.), 1996, vol. 125, pp. 97вАУ107.
- Searle, J.B. and Wojcik, J.M., Chromosomal Evolution: The Case of Sorex araneus, Evolution of Shrews, Wojcik, J.M. and Wolsan, M., Eds., Bialowieza: Mam-mal Res. Inst., Polish Academy of Sciences, 1998, pp. 219вАУ268.
- Halkka, L., Kaikusalo, A., and Vakula, N., Revision of Sorex araneus L. Chromosome Nomenclature, and Race N New to Finland, Ann. Zool. Fennici, 1994, vol. 31, pp. 283вАУ288.
- Fredga, K., The Chromosome Races of Sorex araneus in Scandinavia, Hereditas (Lund, Swed.), 1996, vol. 125, pp. 123вАУ135.
- Searle, J.B., Three New Karyotypic Races of the Com-mon Shrew Sorex araneus (Mammalia, Insectivora) and a Phylogeny, Syst. Zool., 1984, vol. 33, pp. 184вАУ194.
- Wojcik, J.M., Chromosome Races of the Common Shrew Sorex araneus in Poland: A Model of Karyotype Evolution, Acta Theriol., 1993, vol. 38, pp. 315вАУ338.
- Wittenbach, A. and Hauser, J., The Fixation of Metacen-tric Chromosomes during the Chromosomal Evolution in the Common Shrew (Sorex araneus, Insectivora), Hereditas (Lund, Swed.), 1996, vol. 125, pp. 209вАУ217.
- Searle, J.B., Chromosomal Hybrid Zones in Eutherian Mammals, Hybrid Zones and the Evolutionary Process, Harrison, R.G., Ed., New York: Oxford Univ. Press, 1993, pp. 309вАУ353.
- Halkka, L., Soderlund, V., Scaren, U., and Heikkila, J., Chromosomal Polymorphism and Racial Evolution of Sorex araneus, Hereditas (Lund, Swed.), 1987, vol. 106, pp. 257вАУ275.
- Kral, B. and Radjabli, S.I., Banding Patterns and Robert-sonian Fusion in the Western Siberian Population of Sorex araneus (Insectivora, Soricidae), Zool. Listy, 1974,vol. 23, pp. 217вАУ227.
- Aniskin, V.M. and Volobuev, V.T., Chromosome Poly-morphism in Siberian Populations of Shrew of the ara-neusвАУarcticus Complex (Insectivora, Soricidae): III. Three Chromosomal Forms of the Common Shrew Sorex ara-neus L., Genetika (Moscow), 1981, vol. 17, no. 10,pp. 1784вАУ1791.
- Kral, B., Aniskin, V.M., and Volobouev, V.T., Karyotype Variability in Siberian Populations of Sorex araneus (Sori-cidae, Insectivora), Folia Zool., 1981, vol. 30, pp. 23вАУ37.
- Zima, J., Macholan, M., Filippucci, M.G., et al., Karyo-typic and Biochemical Status of Certain Marginal Popu-lations of Sorex araneus, Folia Zool., 1994, vol. 43, suppl. 1, pp. 43вАУ51.
- Polyakov, A.V., Borodin, P.M., Volobuev, V.T., and Searle, J.B., Karyotypic Races of the Common Shrew (Sorex araneus) with Exceptionally Large Ranges: The Novosibirsk and Tomsk Races of Siberia, Hereditas(Lund, Swed.), 1996, vol. 125, pp. 109вАУ115.
- Polyakov, A.V., Chadova, N.B., Rodionova, M.I., et al., Novosibirsk Revisited 24 Years on: Chromosome Poly-morphism in the Novosibirsk Population of the Common Shrew Sorex araneus L., Heredity, 1997, vol. 79, pp. 172вАУ177.
- Polyakov, A.V., Borodin, P.M., Lukacova, L., et al., The Hypothetical Old-Northern Chromosome Race of Sorex araneus Found in the Ural Mountains, Ann. Zool. Fen-nici, 1997, vol. 34, pp. 139вАУ142.
- Polyakov, A.V., Rogacheva, M.B., and Borodin, P.M., Chromosome Geography of Natural Populations of Shrew Sorex araneus and Suncus murinus, Genetika(Moscow), 1997, vol. 33, no. 8, pp. 1126вАУ1132.
- Borodin, P.M., Ladygina, T.Yu., Polyakov, A.V., and Rogacheva, M.B., Chromosome Pairing in Robertsonian Heterozygotes of Common Sorex araneus and Musk Sorex murinus Shrew, Dokl. Ross. Akad. Nauk, 1997, vol. 356, no. 1, pp. 132вАУ134.
- Polyakov, A.V., Zima, J., Searle, J.B., et al., Chromo-some Races of the Common Shrew, Sorex araneus, in the Ural Mountains: A Link between Siberia and Scandina-via?, Acta Theriol., 2000, vol. 45, suppl. 1, pp. 19вАУ26.
- Polyakov, A.V., Zima, J., Banaszek, A., et al., New Chro-mosome Races of the Common Shrew (Sorex araneus) from Eastern Siberia, Acta Theriol., 2000, vol. 45, suppl. 1, pp. 11вАУ18.
- Aniskin, V.M. and LukвАЩyanova, I.V., A New Chromo-some Race and Analysis of the Hybrid Zone of TwoKaryomorphs of Sorex araneus (Insectivora, Soricidae), Dokl. Akad. Nauk SSSR, 1989, vol. 309, pp. 1260вАУ1262.
- Gureev, A.A., Insectivores: Hedgehogs, Moles, and Shrews (Erinaceidae, Talpidae, Soricidae), in Fauna SSSR. Mlekopitayushchie (Fauna of the Soviet Union: Mam-mals), Leningrad: Nauka, 1970, vol. 4, no. 2, p. 503.
- Velichko, A.A., Razvitie landshaftov i klimata severnoi Evrazii (Development of the Landscape and Climate of Northern Eurasia), Moscow: Nauka, 1993, no. 1, p. 102.
- Grichuk, V.P., Main Types of Vegetation (Ecosystems) during the Maximum Cooling of the Last Glaciation, Atlas of Palaeoclimates and Palaeoenvironments of the Northern Hemisphere, Frenzel, B., Pecsi, B., and Veli-chko, A.A., Eds., Budapest: Hung. Acad. Sci., 1992.
- van Campo, E., Guiot, J., and Peng, C., A Data-Based Re-Appraisal of the Terrestrial Carbon Budget at the Last Glacial Maximum, Global and Planetary Change, 1993, vol. 8, pp. 189вАУ201.
- Frenzel, B., Vegetation during the Maximum Cooling of the Last Glaciation, Atlas of Palaeoclimates and Palae-oenvironments of the Northern Hemisphere, Frenzel, B., Pecsi, B., and Velichko, A.A., Eds., Budapest: Hung. Acad. Sci., 1992.
- Baryshnikov, G.F. and Markova, A.K., Main Mammal Assemblages of the Upper Pleniglacial of the Last Gla-ciation, Atlas of Palaeoclimates and Palaeoenviron-ments of the Northern Hemisphere, Frenzel, B., Pecsi, B., and Velichko, A.A., Eds., Budapest: Hung. Acad. Sci., 1992.
- Gropp, A. and Winking, H., Robertsonian Transloca-tions: Cytology, Meiosis, Segregation Patterns and Bio-logical Consequences of Heterozygosity, Symp. Zool. Soc. London, 1981, vol. 47, pp. 141вАУ181.
- Hewitt, G.M., Some Genetic Consequences of Ice Ages, and Their Role in Divergence and Speciation, Biol. J. Linn. Soc., 1996, vol. 58, pp. 247вАУ276.
- Fedyk, S., Banaszek, A., Chetnicki, W., et al., Reassess-ment of the Range of the Drnholec Race: Studies on Meiosis in Sorex araneus Hybrids, Acta Theriol., 2000, vol. 45, suppl. 1, pp. 59вАУ67.
–Ъ–Њ–љ–µ—З–љ–Њ, –і–∞–љ–љ–∞—П —Б—В–∞—В—М—П –љ–µ –±—Л–ї–∞ –љ–∞–њ–Є—Б–∞–љ–∞ —Б–њ–µ—Ж–Є–∞–ї—М–љ–Њ –і–ї—П –Р-—Б–∞–є—В–∞ –Є –∞–≤—В–Њ—А—Л –љ–µ —Б—В–∞–≤–Є–ї–Є —Б–≤–Њ–µ–є –µ–і–Є–љ—Б—В–≤–µ–љ–љ–Њ–є —Ж–µ–ї—М—О-–і–Њ—Б–∞–і–Є—В—М –≤–µ—А—Г—О—Й–Є–Љ.–Ч–∞–Њ—Б—В—А—С–љ–љ—Л–µ –∞–Ї—Ж–µ–љ—В—Л, —Б–≤–Њ–є—Б—В–≤–µ–љ–љ—Л–µ —Б—В–∞—В—М—П–Љ –∞—В–µ–Є—Б—В–Њ–≤, –Ј–і–µ—Б—М –Њ—В—Б—Г—В—Б—В–≤—Г—О—В.–Ю–і–љ–∞–Ї–Њ, –Є–Ј –њ—А–Є–≤–µ–і—С–љ–љ–Њ–≥–Њ –≤ –ґ—Г—А–љ–∞–ї–µ вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ, —Б—В–∞–ї–Є –њ–Њ–љ—П—В–љ—Л –њ—Г—В–Є —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ–≥–Њ —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—П (–љ–∞–Ї–Њ–њ–ї–µ–љ–Є—П) —А–∞–Ј–ї–Є—З–Є–є –≤ –Ї–∞—А–Є–Њ—В–Є–њ–∞—Е —А–∞—Б –±—Г—А–Њ–Ј—Г–±–Ї–Є –Є –Љ–µ—Е–∞–љ–Є–Ј–Љ –і–Њ—Б—В–Є–ґ–µ–љ–Є—П —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є –Љ–µ–ґ—А–∞—Б–Њ–≤—Л—Е –≥–Є–±—А–Є–і–Њ–≤ (–Є–Ј–Њ–ї—П—Ж–Є–Є), –Ї–Њ–≥–і–∞ —А–∞—Б–Њ–≤—Л–µ —А–∞–Ј–ї–Є—З–Є—П —Б—В–∞–љ–Њ–≤—П—В—Б—П —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Л–Љ–Є:
вАЬ–Т –Љ–µ–є–Њ–Ј–µ —Г –≥–Є–±—А–Є–і–Њ–≤ –Љ–µ–ґ–і—Г –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є –Є —В–Њ–Љ—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є –і–Њ–ї–ґ–љ–∞ –Њ–±—А–∞–Ј–Њ–≤—Л–≤–∞—В—М—Б—П —Ж–µ–њ—М –Є–Ј 9 —Н–ї–µ–Љ–µ–љ—В–Њ–≤.–Я–Њ–і–Њ–±–љ—Л–µ –љ–∞—А—Г—И–µ–љ–Є—П, –Ї–∞–Ї –Є–Ј–≤–µ—Б—В–љ–Њ, –і–Њ–ї–ґ–љ—Л –њ—А–Є–≤–Њ–і–Є—В—М –Ї —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О –љ–µ—Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞–љ–љ—Л—Е –≥–∞–Љ–µ—В –Є, –Ї–∞–Ї —Б–ї–µ–і—Б—В–≤–Є–µ –Ї –њ–Њ–ї–љ–Њ–є —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є –Є–ї–Є –Ї —А–µ–Ј–Ї–Њ–Љ—Г —Б–љ–Є–ґ–µ–љ–Є—О –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В–Є –≥–Є–±—А–Є–і–љ—Л—Е –Њ—Б–Њ–±–µ–є [22,29].вАЭ
–Ю—Б—В–∞–≤–∞–ї–Њ—Б—М –љ–µ—П—Б–љ—Л–Љ –і—А—Г–≥–Њ–µ. –Я–Њ —Б—В–∞—В—М–µ —Б–ї–µ–і–Њ–≤–∞–ї–Њ, —З—В–Њ –≤ –±–Њ–ї—М—И–Є–љ—Б—В–≤–µ –і—А—Г–≥–Є—Е –њ–µ—А–µ—Б–µ—З–µ–љ–Є–є –∞—А–µ–∞–ї–Њ–≤ –Њ–±–Є—В–∞–љ–Є—П —А–∞—Б –±—Г—А–Њ–Ј—Г–±–Њ–Ї (–≥–Є–±—А–Є–і–љ—Л—Е –Ј–Њ–љ–∞—Е) –Ї–∞—А–Є–Њ—В–Є–њ–Є—З–µ—Б–Ї–Є–µ –Њ—В–ї–Є—З–Є—П —Б–Ї—А–µ—Й–Є–≤–∞—О—Й–Є—Е—Б—П —А–∞—Б –Љ–Њ–≥—Г—В –±—Л—В—М –Љ–Є–љ–Є–Љ–∞–ї—М–љ—Л–Љ–Є (–Ј–∞—З–∞—Б—В—Г—О –≤ –Њ–і–љ—Г —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є—О –Є–ї–Є —А–Њ–±–µ—А—В—Б–Њ–љ–Њ–≤—Б–Ї–Њ–µ —Б–ї–Є—П–љ–Є–µ).–Ъ–∞–Ї–Њ–≤–∞ —Г –±—Г—А–Њ–Ј—Г–±–Њ–Ї —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Ї –≥–Є–±—А–Є–і–Є–Ј–∞—Ж–Є–Є –Є –љ–∞—Б–Ї–Њ–ї—М–Ї–Њ –њ–ї–Њ–і–Њ–≤–Є—В—Л –≥–Є–±—А–Є–і—Л –≤ —Н—В–Є—Е –Ј–Њ–љ–∞—Е –њ–µ—А–µ—Б–µ—З–µ–љ–Є—П —А–∞—Б (—Б–Љ. —А–Є—Б 2), –≥–і–µ —А–∞–Ј–ї–Є—З–Є—П –Ї–∞—А–Є–Њ—В–Є–њ–Њ–≤ –љ–µ —В–∞–Ї–Є–µ –Ї–Њ–љ—В—А–∞—Б—В–љ—Л–µ (—Б–Љ. —А–Є—Б 1), –Ї–∞–Ї —Г –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є –Є —В–Њ–Љ—Б–Ї–Њ–є —А–∞—Б? –Ъ —Б—З–∞—Б—В—М—О, –≤ —Б–µ—В–Є –љ–∞—И–ї–∞—Б—М –Є –і—А—Г–≥–∞—П —Б—В–∞—В—М—П –Њ–± –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П—Е –≥–Є–±—А–Є–і–љ—Л—Е –Ј–Њ–љ –±—Г—А–Њ–Ј—Г–±–Њ–Ї (–њ–Њ–і —А—Г–Ї–Њ–≤–Њ–і—Б—В–≤–Њ–Љ –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤–∞) , –≥–і–µ –њ–Њ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –Є–Ј—Г—З–µ–љ–Є—П –Ї–∞—А–Є–Њ—В–Є–њ–Њ–≤ –≤—Л—П—Б–љ–Є–ї–Њ—Б—М, —З—В–Њ:
вАЬ–Р—А–µ–∞–ї—Л –і–≤—Г—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л—Е —А–∞—Б –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є —З–∞—Б—В–Є—З–љ–Њ –њ–µ—А–µ–Ї—А—Л–≤–∞—О—В—Б—П –≤ –њ—А–∞–≤–Њ–±–µ—А–µ–ґ—М–µ —А. –Ю–±—М, –Њ–±—А–∞–Ј—Г—П –Ј–Њ–љ—Г, –≥–і–µ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–Є —Н—В–Є—Е —А–∞—Б –≤—Б—В—А–µ—З–∞—О—В—Б—П –Є, —Б–Ї—А–µ—Й–Є–≤–∞—П—Б—М, –Љ–Њ–≥—Г—В –њ—А–Њ–Є–Ј–≤–Њ–і–Є—В—М –Љ–µ–ґ—А–∞—Б–Њ–≤—Л—Е –≥–Є–±—А–Є–і–Њ–≤вАЭ.
–°—Г–і—П –њ–Њ –і–∞–љ–љ—Л–Љ —Б—В–∞—В—М–Є –Є–Ј –ґ—Г—А–љ–∞–ї–∞ вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ, —А–µ—З—М –Є–і—С—В –Њ –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є —А–∞—Б–µ (вДЦ4 –љ–∞ —А–Є—Б.1, —А–Є—Б.2, —В–∞–±–ї–Є—Ж–∞, –ґ-–ї вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ) –Є —Б–Њ—Б–µ–і—Б—В–≤—Г—О—Й–µ–є —Б –љ–µ–є —Б–µ—А–Њ–≤—Б–Ї–Њ–є —А–∞—Б–µ (вДЦ3 –љ–∞ —А–Є—Б. 1, —А–Є—Б.2, —В–∞–±–ї–Є—Ж–∞, –ґ-–ї вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ). –≠—В–Њ —Б–ї–µ–і—Г–µ—В –Є–Ј —В–Њ–≥–Њ, —З—В–Њ –≤ —А–∞–є–Њ–љ–µ —А. –Ю–±—М , –≤ 30-—В–Є –Ї–Љ. –Ј–Њ–љ–µ, –≤–Њ–Ј–ї–µ –≥. –Э–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–∞ –Љ–Њ–≥—Г—В –≥—А–∞–љ–Є—З–Є—В—М —В–Њ–ї—М–Ї–Њ –і–≤–µ —А–∞—Б—Л - —Б–µ—А–Њ–≤—Б–Ї–∞—П –Є –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–∞—П.(—А–Є—Б.2, —В–∞–±–ї–Є—Ж–∞ –≤ –ґ-–ї–µ вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ).–≠—В–Є —А–∞—Б—Л —А–∞–Ј–і–µ–ї—П–µ—В –≤—Б–µ–≥–Њ-–љ–∞–≤—Б–µ–≥–Њ –Њ–і–љ–∞ –њ–Њ–ї–љ–Њ–њ–ї–µ—З–µ–≤–∞—П —А–µ—Ж–Є–њ—А–Њ–Ї–љ–∞—П —В—А–∞–љ—Б–ї–Њ–Ї–∞—Ж–Є—П.–Я–Њ—Б–Ї–Њ–ї—М–Ї—Г –≤ –њ—А–Є–Љ–µ—А–∞—Е —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ–≥–Њ –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –≤–∞–ґ–љ–∞ –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В—М –Є–ї–Є —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В—М –≥–Є–±—А–Є–і–Њ–≤, —В–Њ –≤ –њ–Є—Б—М–Љ–µ —П —Б–њ—А–Њ—Б–Є–ї —Г –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤–∞ –Њ –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В–Є(—Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є) —Б–∞–Љ–Є—Е –≥–Є–±—А–Є–і–Њ–≤ –Є –њ–Њ–ї—Г—З–Є–ї —В–∞–Ї–Њ–є –Њ—В–≤–µ—В :
вАЬ I have to say also that problems with Sorex araneus chromosome races have not yet resolved completely and many things are still in process. However we can say now that hybrids after breeding of two races are fertile, and geographic barriers play non-essential role in isolation of races and supporting of hybrid zonesвАЭ.
(–Ґ–∞–Ї –ґ–µ –і–Њ–ї–ґ–µ–љ —Б–Ї–∞–Ј–∞—В—М, —З—В–Њ –њ—А–Њ–±–ї–µ–Љ—Л –њ–Њ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–Љ —А–∞—Б–∞–Љ –Њ–±—Л–Ї–љ–Њ–≤–µ–љ–љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –і–Њ –Ї–Њ–љ—Ж–∞ –љ–µ —А–µ—И–µ–љ—Л, –Є –Љ–љ–Њ–≥–Є–µ –љ–∞—Е–Њ–і—П—В—Б—П –≤ –њ—А–Њ—Ж–µ—Б—Б–µ(—А–µ—И–µ–љ–Є—П).–Ю–і–љ–∞–Ї–Њ —Б–µ–≥–Њ–і–љ—П –Љ–Њ–ґ–љ–Њ —Б–Ї–∞–Ј–∞—В—М, —З—В–Њ –≥–Є–±—А–Є–і—Л –і–≤—Г—Е —А–∞—Б –њ–ї–Њ–і–Њ–≤–Є—В—Л, –≥–µ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є–µ –±–∞—А—М–µ—А—Л –љ–µ –Є–≥—А–∞—О—В —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–є —А–Њ–ї–Є –≤ –Є–Ј–Њ–ї—П—Ж–Є–Є —А–∞—Б –Є –њ–Њ–і–і–µ—А–ґ–Ї–µ –≥–Є–±—А–Є–і–љ—Л—Е –Ј–Њ–љ.) -
–њ–µ—А–µ–≤–Њ–і –Ь–Њ–Є—Б–µ–µ–љ–Ї–Њ.–Ч–љ–∞—П, –Ї–∞–Ї –Є–љ–Њ–≥–і–∞ "—Б–њ–Њ—А—П—В" –≤–µ—А—Г—О—Й–Є–µ –≤ –і–µ–±–∞—В–љ–µ, —П –Ј–∞–і–∞–ї (–њ–Њ e-mail) —А—П–і –≤–Њ–њ—А–Њ—Б–Њ–≤ —Г—З—С–љ–Њ–Љ—Г –Њ –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В–Є –≥–Є–±—А–Є–і–Њ–≤ –≤ –і—А—Г–≥–Є—Е –Ј–Њ–љ–∞—Е, –≥–і–µ —А–∞—Б—Л –±—Г—А–Њ–Ј—Г–±–Ї–Є –њ–µ—А–µ—Б–µ–Ї–∞—О—В—Б—П. –Я–Њ–љ—П—В–љ–Њ, —З—В–Њ –≤–Њ–њ—А–Њ—Б—Л –Ї–∞—Б–∞–ї–Є—Б—М –≤—Б–µ—Е –Њ—Б—В–∞–ї—М–љ—Л—Е –Є–Ј—Г—З–µ–љ–љ—Л—Е –≥–Є–±—А–Є–і–љ—Л—Е –Ј–Њ–љ –±—Г—А–Њ–Ј—Г–±–Њ–Ї, –≤ –Љ–µ—Б—В–∞—Е –њ–µ—А–µ—Б–µ—З–µ–љ–Є—П –∞—А–µ–∞–ї–Њ–≤ –Є—Е –Њ–±–Є—В–∞–љ–Є—П, –Є—Б–Ї–ї—О—З–∞—П –Ј–Њ–љ—Г —Б—В–µ—А–Є–ї—М–љ—Л—Е –≥–Є–±—А–Є–і–Њ–≤ –Љ–µ–ґ–і—Г –љ–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї–Њ–є –Є —В–Њ–Љ—Б–Ї–Њ–є —А–∞—Б–∞–Љ–Є, –Њ–њ–Є—Б–∞–љ–љ—Г—О (–≤—Л—И–µ) –≤ –ґ—Г—А–љ. вАЬ–У–µ–љ–µ—В–Є–Ї–∞вАЭ.–Э–∞ —Н—В–Њ –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤ –Љ–љ–µ –Њ—В–≤–µ—В–Є–ї:
вАЬThe matter of fact is that in nature chromosome aberrations
(or difference between races)
do not decrease fertility of hybridsвАЭ.
(–£—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –≤ –њ—А–Є—А–Њ–і–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ–љ—Л–µ –∞–±–µ—А—А–∞—Ж–Є–Є (–Є–ї–Є —А–∞–Ј–ї–Є—З–Є—П –Љ–µ–ґ–і—Г —А–∞—Б–∞–Љ–Є) –љ–µ —Г–Љ–µ–љ—М—И–∞—О—В –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В–Є –≥–Є–±—А–Є–і–Њ–≤.)-
–њ–µ—А–µ–≤–Њ–і –Ь–Њ–Є—Б–µ–µ–љ–Ї–Њ–Ф—А—Г–≥–Є–Љ–Є —Б–ї–Њ–≤–∞–Љ–Є, –≤ —В–µ—Е –Њ—Б—В–∞–ї—М–љ—Л—Е –≥–Є–±—А–Є–і–љ—Л—Е –Ј–Њ–љ–∞—Е (–≥–і–µ —А–∞–Ј–ї–Є—З–Є—П –Ї–∞—А–Є–Њ—В–Є–њ–∞ –±—Г—А–Њ–Ј—Г–±–Њ–Ї –љ–µ–±–Њ–ї—М—И–Є–µ) –Њ—В–Љ–µ—З–∞–µ—В—Б—П —Д–∞–Ї—В –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В–Є (–љ–µ —Б—В–µ—А–Є–ї—М–љ–Њ—Б—В–Є) –≥–Є–±—А–Є–і–Њ–≤.
–Э–∞–њ–Њ—Б–ї–µ–і–Њ–Ї —Е–Њ—З—Г –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ –≤—Б–µ –њ—А–Њ—З–Є–µ –≥–Є–њ–Њ—В–µ—В–Є—З–µ—Б–Ї–Є–µ –≤–Њ–њ—А–Њ—Б—Л, –Ї–Њ—В–Њ—А—Л–µ —П –≤ –њ–µ—А–µ–њ–Є—Б–Ї–µ –Ј–∞–і–∞–≤–∞–ї –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤—Г (–∞ —З—В–Њ –±—Г–і–µ—В –µ—Б–ї–Є —В–∞–Ї –Є–ї–Є —Н–і–∞–Ї), –љ–µ –Є–Љ–µ—О—В –љ–∞ —Б–µ–≥–Њ–і–љ—П—И–љ–Є–є –і–µ–љ—М –Њ—В–≤–µ—В–∞.–Т –љ–µ–≤–Њ–ї–µ (–≤ –ї–∞–±–Њ—А–∞—В–Њ—А–Є—П—Е) –±—Г—А–Њ–Ј—Г–±–Њ–Ї –љ–µ —Б–Њ–і–µ—А–ґ–∞—В –Є —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ –±–Њ–ї–µ–µ —В–Њ–љ–Ї–Є—Е —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–Њ–≤ —Б –і–ї–Є—В–µ–ї—М–љ—Л–Љ –љ–∞–±–ї—О–і–µ–љ–Є–µ–Љ –љ–µ –±—Л–ї–Њ.–≠—В–Њ —В—А–µ–±—Г–µ—В –љ–µ–Љ–∞–ї—Л—Е —Б—А–µ–і—Б—В–≤.–£—З—С–љ—Л–µ –≤—Л–µ–Ј–ґ–∞—О—В –≤ —Н–Ї—Б–њ–µ–і–Є—Ж–Є–Є –Є –Є–Ј—Г—З–∞—О—В –±—Г—А–Њ–Ј—Г–±–Њ–Ї –љ–∞ –Љ–µ—Б—В–µ, –≤ –њ–Њ–ї–µ–≤—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е.–≠—В–Њ –≤—Б—С, —З—В–Њ –љ–∞ —Б–µ–≥–Њ–і–љ—П –њ—А–Њ–і–µ–ї–∞–љ–Њ.–Т –Њ–і–љ–Њ–Љ –Є–Ј –њ–Њ—Б–ї–µ–і–љ–Є—Е –њ–Є—Б–µ–Љ –Р.–Т.–Я–Њ–ї—П–Ї–Њ–≤ –Љ–љ–µ —В–∞–Ї –Є –љ–∞–њ–Є—Б–∞–ї:
вАЬProbably (and very probable) after the summer seazon of this
year, which we are going to
spend on hybrid zones, we will publicate the answer on this questionвАЭ.
(–Т–Њ–Ј–Љ–Њ–ґ–љ–Њ (–Є –Њ—З–µ–љ—М –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ), —З—В–Њ –њ–Њ—Б–ї–µ –ї–µ—В–љ–µ–≥–Њ —Б–µ–Ј–Њ–љ–∞ –≤ —Н—В–Њ–Љ –≥–Њ–і—Г(2002), –Ї–Њ—В–Њ—А—Л–є –Љ—Л –њ–ї–∞–љ–Є—А—Г–µ–Љ –њ—А–Њ–≤–µ—Б—В–Є –≤ –≥–Є–±—А–Є–і–љ—Л—Е –Ј–Њ–љ–∞—Е, –Љ—Л –Њ–њ—Г–±–ї–Є–Ї—Г–µ–Љ –Њ—В–≤–µ—В—Л –љ–∞ —Н—В–Є –≤–Њ–њ—А–Њ—Б—Л).-
–њ–µ—А–µ–≤–Њ–і –° –Ь–Њ–Є—Б–µ–µ–љ–Ї–Њ.–Ф—Г–Љ–∞—О, —З—В–Њ –њ—А–Њ–і–µ–ї–∞–љ–љ–Њ–є –љ–∞ —Б–µ–≥–Њ–і–љ—П —А–∞–±–Њ—В—Л, –њ–Њ –Є–Ј—Г—З–µ–љ–Є—О –±—Г—А–Њ–Ј—Г–±–Њ–Ї, –≤–Њ–≤—Б–µ –љ–µ –Љ–∞–ї–Њ, –і–ї—П —Г–±–µ–і–Є—В–µ–ї—М–љ–Њ–≥–Њ –Є –љ–∞–±–ї—О–і–∞–µ–Љ–Њ–≥–Њ –≤ –њ—А–Є—А–Њ–і–µ –њ—А–Є–Љ–µ—А–∞ –Њ–і–љ–Њ–≥–Њ –Є–Ј —Н—В–∞–њ–Њ–≤ –≤–Є–і–Њ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П.
***
–Ъ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Г—З—С–љ—Л—Е –љ–µ—В —Б–Љ—Л—Б–ї–∞ –і–Њ–±–∞–≤–ї—П—В—М –Њ—В—Б–µ–±—П—В–Є–љ—Г.–Ф–ї—П –њ–Њ–ї–љ–Њ—В—Л –Ї–∞—А—В–Є–љ—Л –њ—А–Є–≤–µ–і—Г –љ–µ–Ї–Њ—В–Њ—А—Л–µ –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ—Л–µ —Б–≤–µ–і–µ–љ–Є—П –Є–Ј –±–Є–Њ–ї–Њ–≥–Є–Є, –Ї–Њ—В–Њ—А—Л–µ –Ї–∞—Б–∞—О—В—Б—П –±—Г—А–Њ–Ј—Г–±–Њ–Ї.
–С—Г—А–Њ–Ј—Г–±–Ї–Є, —А–Њ–і –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, —Б–µ–Љ–µ–є—Б—В–≤–Њ –Ј–µ–Љ–ї–µ—А–Њ–µ–Ї, –Ф–ї–Є–љ–∞ —В–µ–ї–∞ 4-9 —Б–Љ.,—Е–≤–Њ—Б—В–∞ 2.5-8 —Б–Љ.,–Ъ–Њ—А–Њ–љ–Ї–Є –Ј—Г–±–Њ–≤ –Ї—А–∞—Б–љ–Њ–≤–∞—В—Л –±—Г—А—Л–µ 50-65 –≤–Є–і–Њ–≤, –≤ –Х–≤—А–∞–Ј–Є–Є, –°–µ–≤ –Р–Љ–µ—А–Є–Ї–µ, –Є —Б–µ–≤ —З–∞—Б—В–Є –Ѓ–ґ–љ.–Р–Љ–µ—А–Є–Ї–Є. –Т –°–°–°–† 16-18 –≤–Є–і–Њ–≤. –С. -–Ї—А–Њ—И–Ї–∞, –Њ–і–љ–Њ –Є–Ј —Б–∞–Љ—Л—Е –Љ–∞–ї–µ–љ—М–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –≤–µ—Б–Є—В –Њ–Ї 2 –≥—А., 2 –≤–Є–і–∞ –≤ –Ї—А–∞—Б–љ–Њ–є –Ї–љ–Є–≥–µ –°–°–°–†.
(–°–Њ–≤.—Н–љ—Ж. —Б–ї–Њ–≤–∞—А—М –њ–Њ–і —А–µ–і. –Я—А–Њ—Е–Њ—А–Њ–≤–∞ 1989 –≥.).
–Я–Њ –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є–Є –±—Г—А–Њ–Ј—Г–±–Ї–∞ –Њ—В–љ–Њ—Б–Є—В—Б—П –Ї –њ–Њ–і–Ї–ї–∞—Б—Б—Г –њ–ї–∞—Ж–µ–љ—В–∞—А–љ—Л—Е ( —З—В–Њ –Є —З–µ–ї–Њ–≤–µ–Ї ), –њ—А–∞–≤–і–∞ –Ї –і—А—Г–≥–Њ–Љ—Г –Њ—В—А—П–і—Г - –љ–∞—Б–µ–Ї–Њ–Љ–Њ—П–і–љ—Л–Љ (—З–µ–ї–Њ–≤–µ–Ї –≤ –Њ—В—А—П–і–µ –њ—А–Є–Љ–∞—В–Њ–≤).
–Я–ї–∞—Ж–µ–љ—В–∞—А–љ—Л–µ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є–µ - —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є–µ–Љ –њ–ї–∞—Ж–µ–љ—В—Л , —З–µ—А–µ–Ј –Ї–Њ—В–Њ—А—Г—О –њ–Є—В–∞–µ—В—Б—П —Н–Љ–±—А–Є–Њ–љ –≤ –њ–µ—А–Є–Њ–і –≤–љ—Г—В—А–Є—Г—В—А–Њ–±–љ–Њ–≥–Њ —А–∞–Ј–≤–Є—В–Є—П.–Я–ї–∞—Ж–µ–љ—В–∞ –Њ–±—А–∞–Ј—Г–µ—В—Б—П —З–∞—Б—В—М—О –Є–Ј —В–Ї–∞–љ–µ–є —Н–Љ–±—А–Є–Њ–љ–∞–ї—М–љ–Њ–≥–Њ –њ—А–Њ–Є—Б—Е–Њ–ґ–і–љ–µ–љ–Є—П, —З–∞—Б—В—М—О –Є–Ј —В–Ї–∞–љ–µ–є –Љ–∞—В–µ—А–Є; –њ—А–Є –њ–Њ–Љ–Њ—Й–Є –њ–ї–∞—Ж–µ–љ—В—Л —Н–Љ–±—А–Є–Њ–љ –њ–Њ–ї—Г—З–∞–µ—В –њ–Є—В–∞—В–µ–ї—М–љ—Л–µ –≤–µ—Й–µ—Б—В–≤–∞ –Є –Ї–Є—Б–ї–Њ—А–Њ–і –Є –Њ—Б–≤–Њ–±–Њ–ґ–і–∞–µ—В—Б—П –Њ—В –њ—А–Њ–і—Г–Ї—В–Њ–≤ –Њ–±–Љ–µ–љ–∞.–Ф–µ—В—С–љ—Л—И–Є —А–Њ–ґ–і–∞—О—В—Б—П –ґ–Є–≤—Л–Љ–Є; –≤ –Љ–Њ–Љ–µ–љ—В —А–Њ–ґ–і–µ–љ–Є—П –Њ–љ–Є –±–Њ–ї–µ–µ —А–∞–Ј–≤–Є—В—Л, —З–µ–Љ –љ–Њ–≤–Њ—А–Њ–ґ–і—С–љ–љ—Л–µ —Б—Г–Љ—З–∞—В—Л–µ (–њ–Њ–і–Ї–ї–∞—Б—Б –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е).
("–С–Є–Њ–ї–Њ–≥–Є—П", –Т–Є–ї–ї–Є, –Ь–Є—А, 1964–≥., —Б—В—А 262-263).
–Ц–Є–≤—Г—В –±—Г—А–Њ–Ј—Г–±–Ї–Є –≥–Њ–і, –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ 15-16 –Љ–µ—Б—П—Ж–µ–≤ (–Ц–Є–≤–Њ—В–љ—Л–µ –Ъ—А–∞—Б–љ–Њ—П—А—Б–Ї–Њ–≥–Њ –Ї—А–∞—П) –Є —Б–Љ–µ–љ–∞ –њ–Њ–Ї–Њ–ї–µ–љ–Є–є –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –µ–ґ–µ–≥–Њ–і–љ–Њ (–њ—А–Є–Љ–µ—А–љ–Њ –≤ 20 —А–∞–Ј –±—Л—Б—В—А–µ–µ, —З–µ–Љ —Г —З–µ–ї–Њ–≤–µ–Ї–∞). –Я—А–Є–Ї–Є–љ—Г–≤ —З–Є—Б–ї–Њ –ї–µ—В –і–Њ –ї–µ–і–љ–Є–Ї–Њ–≤–Њ–≥–Њ –њ–µ—А–Є–Њ–і–∞ –Љ–Њ–ґ–љ–Њ –Њ—Ж–µ–љ–Є—В—М —З–Є—Б–ї–Њ —Б–Љ–µ–љ–Є–≤—И–Є—Е—Б—П –њ–Њ–Ї–Њ–ї–µ–љ–Є–є –Є —Б–Ї–Њ—А–Њ—Б—В—М (–≤ –≥–Њ–і–∞—Е) —Н–≤–Њ–ї—О—Ж–Є–Є –±—Г—А–Њ–Ј—Г–±–Њ–Ї.
–Ч–Є–Љ–Њ–є –±—Г—А–Њ–Ј—Г–±–Ї–Є –љ–µ –≤–њ–∞–і–∞—О—В –≤ —Б–њ—П—З–Ї—Г (–≤ —Е–Њ–ї–Њ–і–љ—Л—Е —А–∞–є–Њ–љ–∞—Е —Б –Љ–Њ—А–Њ–Ј–∞–Љ–Є –Є —Б–љ–µ–≥–∞–Љ–Є), –Њ–љ–Є –њ—А–Њ–і–Њ–ї–ґ–∞—О—В –±–Њ–і—А—Б—В–≤–Њ–≤–∞—В—М.
("–Ц–Є–Ј–љ—М –љ–∞ —Б–љ–µ–≥—Г –Є –њ–Њ–і —Б–љ–µ–≥–Њ–Љ", –њ—А–Њ—Д.–У..–Р.–Э–Њ–≤–Є–Ї–Њ–≤, –Ы–µ–љ–Є–љ–≥—А–∞–і, 1981, –Є–Ј–і-–≤–Њ –Ы–У–£ –Є–Љ. –Ц–і–∞–љ–Њ–≤–∞).
–§–Є–Ј–Є–Њ–ї–Њ–≥–Є—П –±—Г—А–Њ–Ј—Г–±–Ї–Є —Б—Е–Њ–і–љ–∞ —Б —З–µ–ї–Њ–≤–µ—З–µ—Б–Ї–Њ–є.–Ъ–∞–Ї –Є –≤—Б–µ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є–µ-–љ–∞–Є–±–Њ–ї–µ–µ –Њ—А–≥–∞–љ–Є–Ј–Њ–≤–∞–љ–љ—Л–є –Ї–ї–∞—Б—Б –њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е - –±—Г—А–Њ–Ј—Г–±–Ї–∞ –Є–Љ–µ–µ—В (–≤—Л–±–Њ—А–Њ—З–љ–Њ) :
1.–Т—Л—Б–Њ–Ї–Њ–µ —А–∞–Ј–≤–Є—В–Є–µ –¶–Э–°
2.–Ц–Є–≤–Њ—А–Њ–ґ–і–µ–љ–Є–µ –Є –≤—Л–Ї–∞—А–Љ–ї–Є–≤–∞–љ–Є–µ –і–µ—В—С–љ—Л—И–µ–є –Љ–Њ–ї–Њ–Ї–Њ–Љ
3.–Т—Л—Б–Њ–Ї–Њ—А–∞–Ј–≤–Є—В—Г—О —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Ї —В–µ—А–Љ–Њ—А–µ–≥—Г–ї—П—Ж–Є–Є (–њ–Њ—Б—В–Њ—П–љ–љ—Г—О —В–µ–Љ–њ–µ—А–∞—В. —В–µ–ї–∞)
–Ь–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є :
1.–Ґ–µ–ї–Њ –њ–Њ–Ї—А—Л—В–Њ —И–µ—А—Б—В—М—О
2.–Ъ–Њ–ґ–∞ –±–Њ–≥–∞—В–∞ –ґ–µ–ї–µ–Ј–∞–Љ–Є, –љ–∞–ї–Є—З–Є–µ –Љ–ї–µ—З–љ—Л—Е –ґ–µ–ї–µ–Ј.
3.–Э–Є–ґ–љ—П—П —З–µ–ї—О—Б—В—М —Б–Њ—Б—В–Њ–Є—В —В–Њ–ї—М–Ї–Њ –Є–Ј –Ј—Г–±–љ–Њ–є –Ї–Њ—Б—В–Є. –∞ –Ї–≤–∞–і—А–∞—В–љ–∞—П –Є —Б–Њ—З–ї–µ–љ–Њ–≤–∞—П –Ї–Њ—Б—В–Є –њ—А–µ–≤—А–∞—Й–∞—О—В—Б—П –≤ —Б–ї—Г—Е–Њ–≤—Л–µ –Ї–Њ—Б—В–Њ—З–Ї–Є.
4.–Ч—Г–±—Л –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ—Л –љ–∞ —А–µ–Ј—Ж—Л. –Ї–ї—Л–Ї–Є –Є –Ї–Њ—А–µ–љ–љ—Л–µ.
5.–°–µ—А–і—Ж–µ —З–µ—В—Л—А—С—Е–Ї–∞–Љ–µ—А–љ–Њ–µ, —Б–Њ—Е—А–∞–љ—П–µ—В—Б—П –Њ–і–љ–∞ –ї–µ–≤–∞—П –і—Г–≥–∞ –∞–Њ—А—В—Л.
6.–≠—А–Є—В—А–Њ—Ж–Є—В—Л –±–µ–Ј—К—П–і–µ—А–љ—Л–µ.
–Т–љ–µ—И–љ–µ–µ —Б—В—А–Њ–µ–љ–Є–µ - —В–µ–ї–Њ –њ–Њ–і—А–∞–Ј–і–µ–ї—П–µ—В—Б—П –љ–∞ –≥–Њ–ї–Њ–≤—Г, —И–µ—О, —В—Г–ї–Њ–≤–Є—Й–µ –Є —Е–≤–Њ—Б—В.
–Я–Њ–Ї—А–Њ–≤( –Ї–Њ–ґ–љ—Л–є) - –±–Њ–ї–µ–µ —Б–ї–Њ–ґ–љ—Л–є, —З–µ–Љ —Г –і—А—Г–≥–Є—Е –њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е.
–°–Ї–µ–ї–µ—В –Є –Љ—Г—Б–Ї—Г–ї–∞—В—Г—А–∞ - —Б–Њ—Б—В–Њ–Є—В –Є–Ј —Б–Ї–µ–ї–µ—В–∞ –≥–Њ–ї–Њ–≤—Л, –њ–Њ–Ј–≤–Њ–љ–Њ—З–љ–Є–Ї–∞,, –≥—А—Г–і–љ–Њ–є –Ї–ї–µ—В–Ї–Є, –њ–Њ—П—Б–∞ –њ–µ—А–µ–і–љ–Є—Е –Є –Ј–∞–і–љ–Є—Е –Ї–Њ–љ–µ—З–љ–Њ—Б—В–µ–є –Є —Б–≤–Њ–±–Њ–і–љ—Л—Е –Ї–Њ–љ–µ—З–љ–Њ—Б—В–µ–є.
–Ь—Л—И–µ—З–љ–∞—П —Б–Є—Б—В–µ–Љ–∞ –і–Њ—Б—В–Є–≥–∞–µ—В –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ–≥–Њ —А–∞–Ј–≤–Є—В–Є—П –Є —Б–ї–Њ–ґ–љ–Њ—Б—В–Є, –љ–∞—Б—З–Є—В—Л–≤–∞–µ—В –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —Б–Њ—В –Љ—Л—И—Ж.
–Я–Є—Й–µ–≤–∞—А–Є—В–µ–ї—М–љ–∞—П —Б–Є—Б—В–µ–Љ–∞ - —Б–ї–Њ–ґ–љ–∞—П, —А–∞–Ј–≤–Є—В—Л –њ–Є—Й–µ–≤–∞—А–Є—В. –ґ–µ–ї–µ–Ј—Л.(—Б–ї—О–љ–љ—Л–µ, –њ–Њ–і–ґ–µ–ї—Г–і–Њ—З–љ–∞—П, –њ–µ—З–µ–љ—М).
–Ф—Л—Е–∞—В–µ–ї—М–љ–∞—П —Б–Є—Б—В–µ–Љ–∞ - –ї–µ–≥–Ї–Є–µ –Є —В–Њ–ї—М–Ї–Њ 1% —З–µ—А–µ–Ј –Ї–Њ–ґ–љ—Л–є –њ–Њ–Ї—А–Њ–≤.
–Ъ—А–Њ–≤–µ–љ–Њ—Б–љ–∞—П —Б–Є—Б—В–µ–Љ–∞ - –і–Њ—Б—В–∞–≤–ї—П–µ—В –Ї–Є—Б–ї–Њ—А–Њ–і –Є –њ–Є—В–∞—В. –≤–µ—Й–µ—Б—В–≤–∞ –Ї —В–Ї–∞–љ—П–Љ.
–Т—Л–і–µ–ї–Є—В–µ–ї—М–љ. —Б–Є—Б—В–µ–Љ–∞ - –њ–Њ—З–Ї–Є, –Љ–Њ—З–µ–≤–Њ–є –њ—Г–Ј—Л—А—М, –њ–Њ—В–Њ–≤—Л–µ –ґ–µ–ї–µ–Ј—Л.
–Э–µ—А–≤–љ–∞—П —Б–Є—Б—В–µ–Љ–∞ - –љ–∞–Є–±–Њ–ї–µ–µ –Њ—А–≥–∞–љ–Є–Ј–Њ–≤–∞–љ–∞—П —Г –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е.
–†–∞–Ј–Љ–љ–Њ–ґ–µ–љ–Є–µ –Є —А–∞–Ј–≤–Є—В–Є–µ - –њ–Њ–ї–Њ–≤–Њ–µ —Г —Б–∞–Љ–Њ–Ї –њ–∞—А–љ—Л–µ —П–Є—З–љ–Є–Ї–Є, —П–є—Ж–µ–≤–Њ–і—Л, –Љ–∞—В–Ї–∞ , –≤–ї–∞–≥–∞–ї–Є—Й–µ, –њ—А–µ–і–≤–µ—А–Є–µ –≤–ї–∞–≥–∞–ї–Є—Й–∞.–£ —Б–∞–Љ—Ж–Њ–≤ –њ–∞—А–љ—Л–µ —Б–µ–Љ–µ–љ–љ–Є–Ї–Є, —Б–µ–Љ—П–њ—А–Њ–≤–Њ–і, –Љ–Њ—З–µ–њ–Њ–ї–Њ–≤–Њ–є –Ї–∞–љ–∞–ї –њ–Њ–ї–Њ–≤–Њ–≥–Њ —З–ї–µ–љ–∞.
–Я–Њ—Б–ї–µ —А–Њ–ґ–і–µ–љ–Є—П –і–µ—В—С–љ—Л—И –њ–Є—В–∞–µ—В—Б—П –Љ–Њ–ї–Њ–Ї–Њ–Љ –Љ–∞—В–µ—А–Є, –њ–Њ—Б–ї–µ –≤—Б–Ї–∞—А–Љ–ї–Є–≤–∞–љ–Є—П –Љ–Њ–ї–Њ–Ї–Њ–Љ —Б–≤—П–Ј—М —А–Њ–і–Є—В–µ–ї–µ–є –Є –њ–Њ—В–Њ–Љ—Б—В–≤–∞ —Б–Њ—Е—А–∞–љ—П–µ—В—Б—П –µ—Й—С –љ–µ–Ї–Њ—В. –≤—А–µ–Љ—П. –і–ї—П –њ–µ—А–µ–і–∞—З–Є –ґ–Є–Ј–љ–µ–љ–љ. –Њ–њ—Л—В–∞ –њ–Њ—В–Њ–Љ—Б—В–≤—Г.
("–С–Є–Њ–ї–Њ–≥–Є—П", –Ш–ґ–µ–≤—Б–Ї, 1995–≥, –£–і–Љ—Г—А—В—Б–Ї–Є–є —Г–љ–Є–≤–µ—А—Б–Є—В–µ—В, —Б—В—А. 129-133) :
–Т–Њ—В –µ—Й—С –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —Б—Б—Л–ї–Њ–Ї –њ–Њ —В–µ–Љ–µ:
–Ц–Є–≤–Њ—В–љ—Л–µ –Ъ—А–∞—Б–љ–Њ—П—А—Б–Ї–Њ–≥–Њ –Ї—А–∞—П
–≠–Т–Ю–Ы–Ѓ–¶–Ш–ѓ –•–†–Ю–Ь–Ю–°–Ю–Ь –Ю–С–Ђ–Ъ–Э–Ю–Т–Х–Э–Э–Ю–Щ –С–£–†–Ю–Ч–£–С–Ъ–Ш
–Ъ—А–Њ—И–µ—З–љ–∞—П –±—Г—А–Њ–Ј—Г–±–Ї–∞
–Я—А–Є–ї–Њ–ґ–µ–љ–Є–µ 1. Russian Journal of Genetics, Vol 37, No 4, 2001,
pp.351-357.
08.02.2002–≥.
–°.–Ь–Њ–Є—Б–µ–µ–љ–Ї–Њ.
E-mail sergosergo@mtu-net.ru
–Ъ–Њ–њ–Є—А–Њ–≤–∞–љ–Є–µ, —В–Є—А–∞–ґ–Є—А–Њ–≤–∞–љ–Є–µ,
–њ–µ—А–µ–њ–µ—З–∞—В–Ї–∞, —Б—Б—Л–ї–Ї–∞ –Є –њ—А–Њ—З–Є–µ —Б–њ–Њ—Б–Њ–±—Л
—А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є—П —Н—В–Њ–є —Б—В–∞—В—М–Є –њ—А–Є–≤–µ—В—Б—В–≤—Г—О—В—Б—П –Є
–Њ–і–Њ–±—А—П—О—В—Б—П –∞–≤—В–Њ—А–Њ–Љ –±–µ–Ј –≤—Б—П–Ї–Є—Е –њ—А–µ—В–µ–љ–Ј–Є–є.